

ATM is required for efficient recombination between immunoglobulin switch regions
- PMID: 15520243
- PMCID: PMC2211855
- DOI: 10.1084/jem.20041162
ATM is required for efficient recombination between immunoglobulin switch regions
Abstract
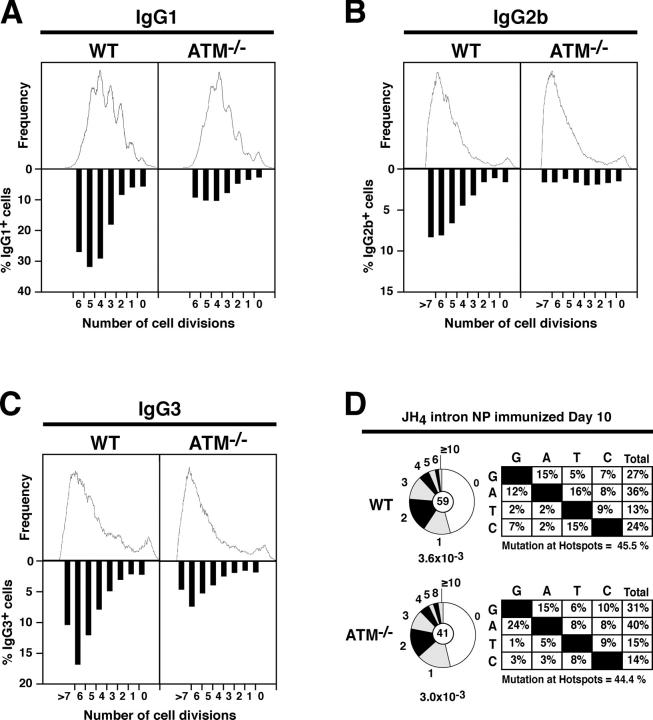
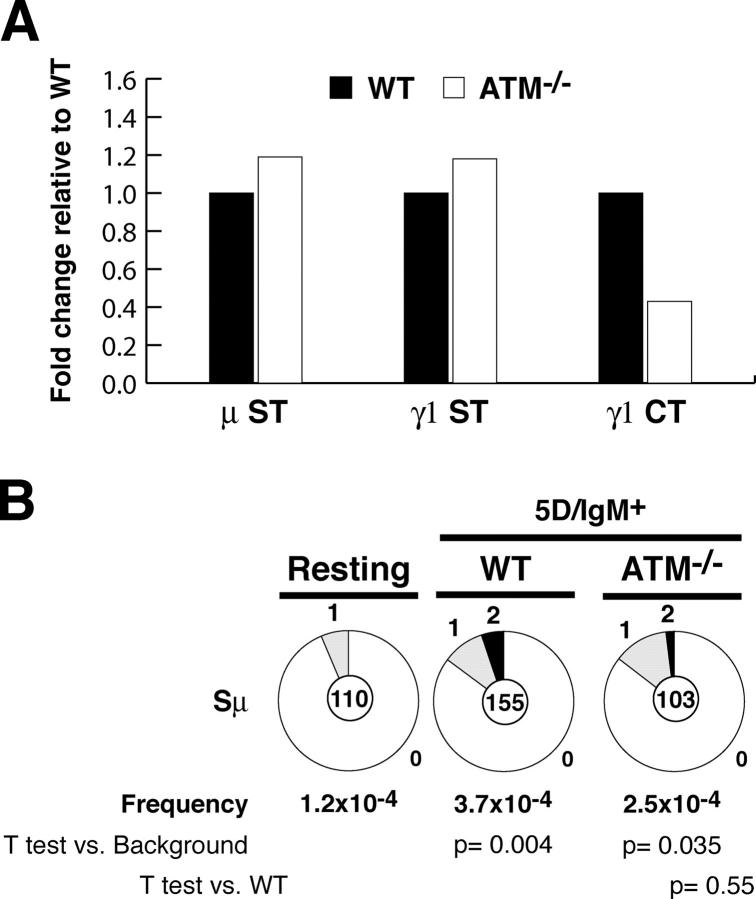
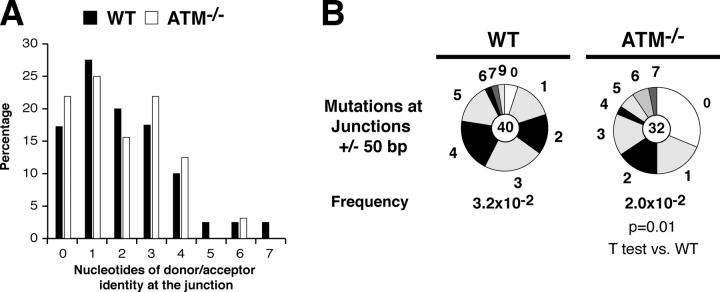
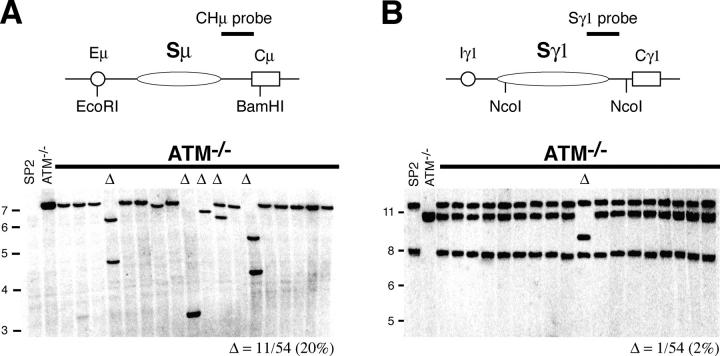
Ataxia telangiectasia mutated (ATM) kinase is critical for initiating the signaling pathways that lead to cell cycle checkpoints and DNA double strand break repair. In the absence of ATM, humans and mice show a primary immunodeficiency that includes low serum antibody titers, but the role of ATM in antigen-driven immunoglobulin gene diversification has not been defined. Here, we show that although ATM is dispensable for somatic hypermutation, it is required for efficient class switch recombination (CSR). The defect in CSR is not due to alterations in switch region transcription, accessibility, DNA damage checkpoint protein recruitment, or short-range intra-switch region recombination. Only long-range inter-switch recombination is defective, indicating an unexpected role for ATM in switch region synapsis during CSR.
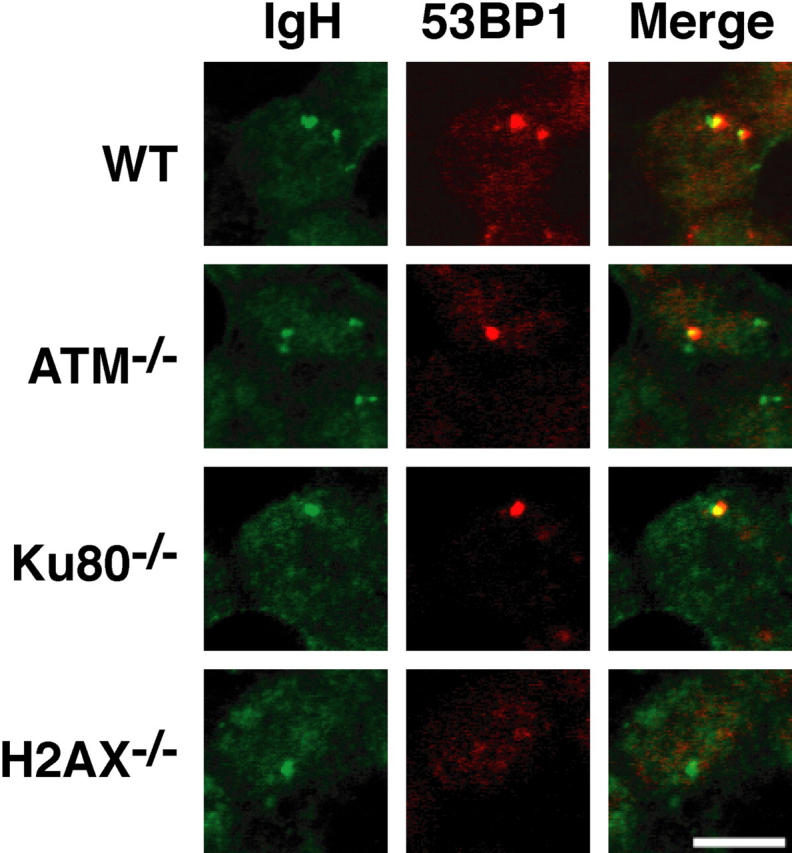
Figures
References
-
- Stavnezer, J. 1996. Immunoglobulin class switching. Curr. Opin. Immunol. 8:199–205. - PubMed
-
- Manis, J.P., M. Tian, and F.W. Alt. 2002. Mechanism and control of class-switch recombination. Trends Immunol. 23:31–39. - PubMed
-
- Muramatsu, M., K. Kinoshita, S. Fagarasan, S. Yamada, Y. Shinkai, and T. Honjo. 2000. Class switch recombination and hypermutation require activation-induced cytidine deaminase (AID), a potential RNA editing enzyme. Cell. 102:553–563. - PubMed
-
- Revy, P., T. Muto, Y. Levy, F. Geissmann, A. Plebani, O. Sanal, N. Catalan, M. Forveille, R. Dufourcq-Labelouse, A. Gennery, et al. 2000. Activation-induced cytidine deaminase (AID) deficiency causes the autosomal recessive form of the Hyper-IgM syndrome (HIGM2). Cell. 102:565–575. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
