

Mitochondrial fission proteins regulate programmed cell death in yeast
- PMID: 15520274
- PMCID: PMC528898
- DOI: 10.1101/gad.1247904
Mitochondrial fission proteins regulate programmed cell death in yeast
Abstract
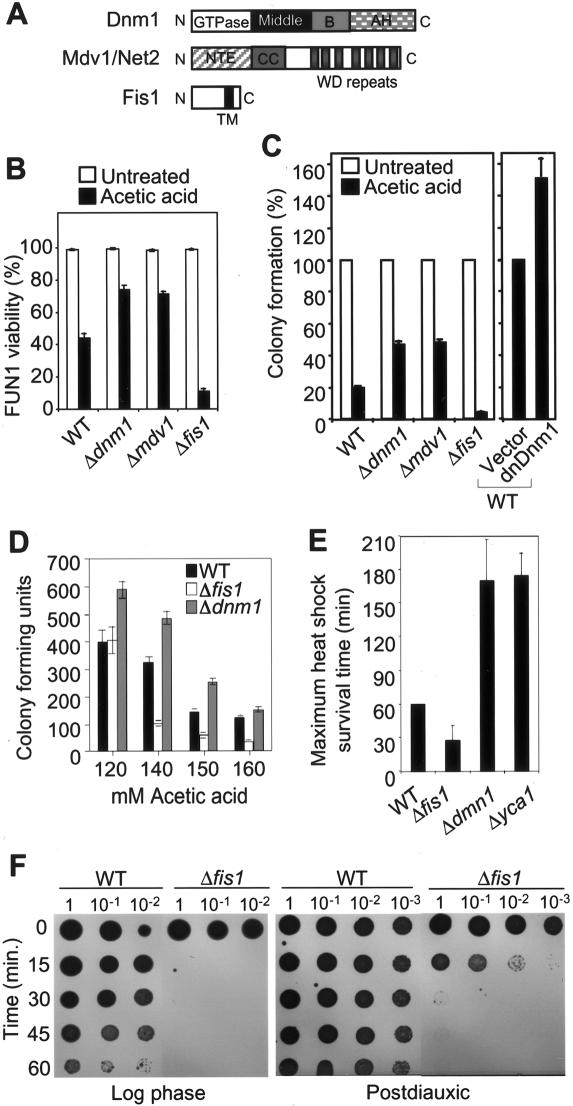
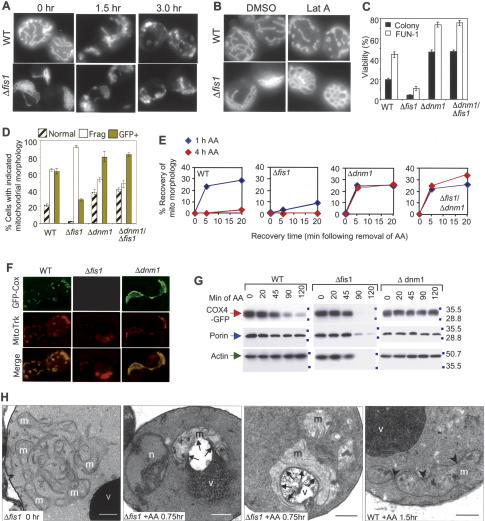
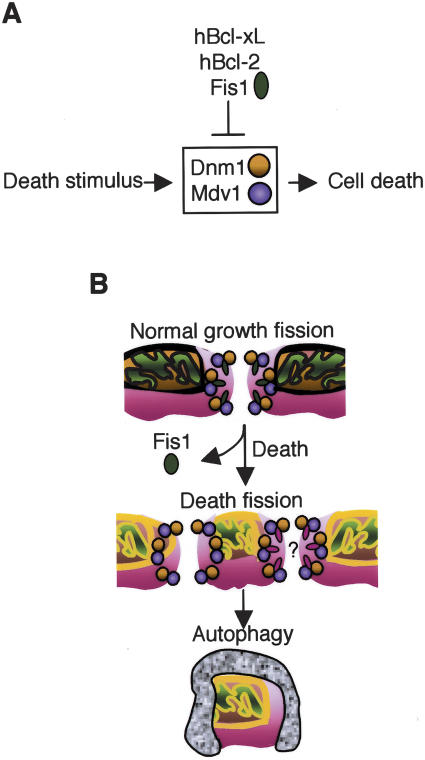
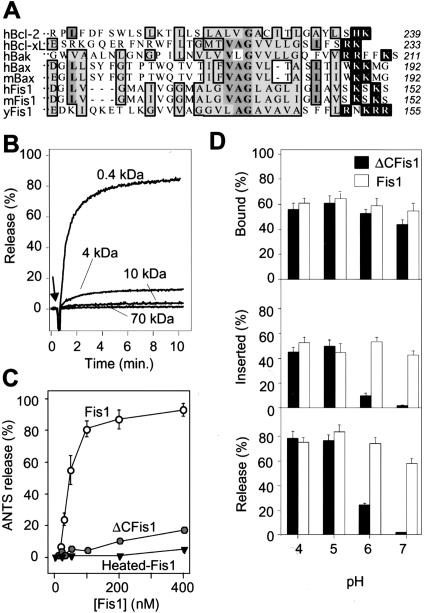
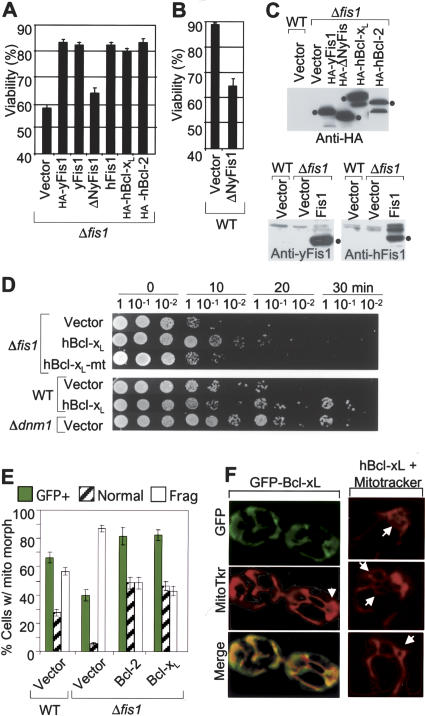
The possibility that single-cell organisms undergo programmed cell death has been questioned in part because they lack several key components of the mammalian cell death machinery. However, yeast encode a homolog of human Drp1, a mitochondrial fission protein that was shown previously to promote mammalian cell death and the excessive mitochondrial fragmentation characteristic of apoptotic mammalian cells. In support of a primordial origin of programmed cell death involving mitochondria, we found that the Saccharomyces cerevisiae homolog of human Drp1, Dnm1, promotes mitochondrial fragmentation/degradation and cell death following treatment with several death stimuli. Two Dnm1-interacting factors also regulate yeast cell death. The WD40 repeat protein Mdv1/Net2 promotes cell death, consistent with its role in mitochondrial fission. In contrast to its fission function in healthy cells, Fis1 unexpectedly inhibits Dnm1-mediated mitochondrial fission and cysteine protease-dependent cell death in yeast. Furthermore, the ability of yeast Fis1 to inhibit mitochondrial fission and cell death can be functionally replaced by human Bcl-2 and Bcl-xL. Together, these findings indicate that yeast and mammalian cells have a conserved programmed death pathway regulated by a common molecular component, Drp1/Dnm1, that is inhibited by a Bcl-2-like function.
Figures


References
-
- Amarante-Mendes G.P., Finucane, D.M., Martin, S.J., Cotter, T.G., Salvesen, G.S., and Green, D.R. 1998. Anti-apoptotic oncogenes prevent caspase-dependent and independent commitment for cell death. Cell Death Differ. 5: 298-306. - PubMed
-
- Basañez G., Zhang, J., Chau, B.N., Maksaev, G.I., Frolov, V., Brandt, T.A., Burch, J., Hardwick, J.M., and Zimmerberg, J. 2001. Pro-apoptotic cleavage products of Bcl-xL form cytochrome c-conducting pores in pure lipid bilayers. J. Biol. Chem. 276: 31083-31091. - PubMed
-
- Brunet C.L., Gunby, R.H., Benson, R.S., Hickman, J.A., Watson, A.J., and Brady, G. 1998. Commitment to cell death measured by loss of clonogenicity is separable from the appearance of apoptotic markers. Cell Death Differ. 5: 107-115. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous