

The Polycomb Ezh2 methyltransferase regulates muscle gene expression and skeletal muscle differentiation
- PMID: 15520282
- PMCID: PMC525543
- DOI: 10.1101/gad.1241904
The Polycomb Ezh2 methyltransferase regulates muscle gene expression and skeletal muscle differentiation
Erratum in
- Genes Dev. 2005 Mar 15;19(6):768
Abstract
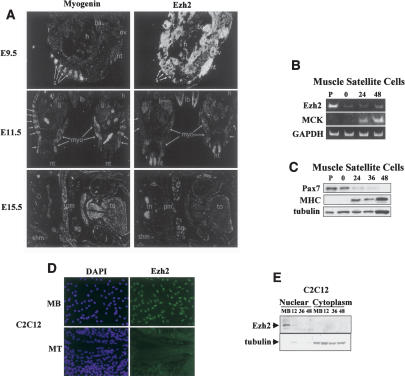
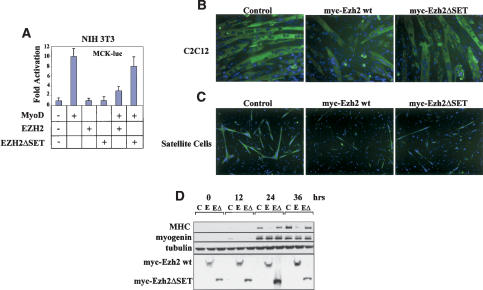
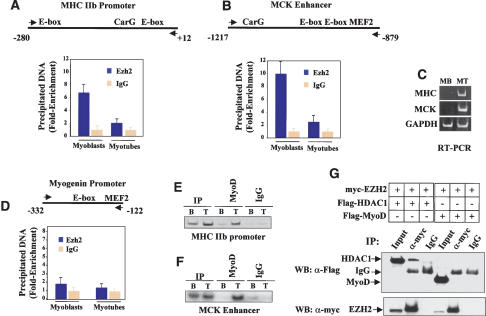
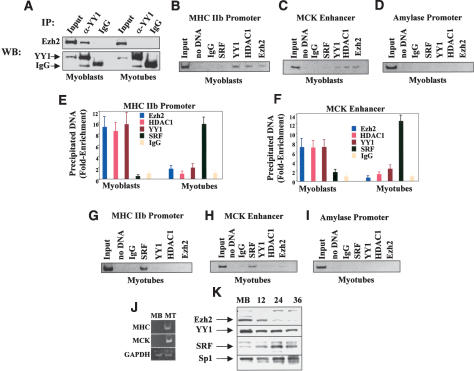
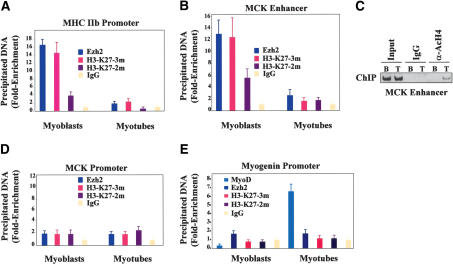
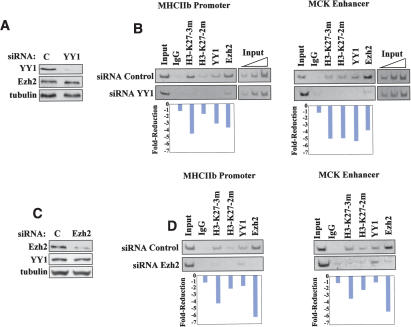
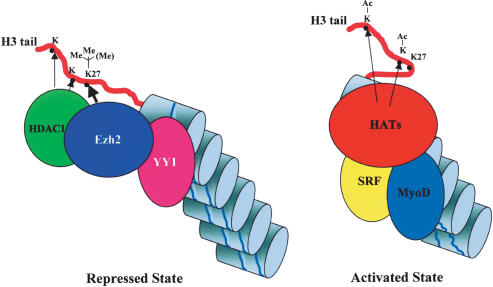
The Ezh2 protein endows the Polycomb PRC2 and PRC3 complexes with histone lysine methyltransferase (HKMT) activity that is associated with transcriptional repression. We report that Ezh2 expression was developmentally regulated in the myotome compartment of mouse somites and that its down-regulation coincided with activation of muscle gene expression and differentiation of satellite-cell-derived myoblasts. Increased Ezh2 expression inhibited muscle differentiation, and this property was conferred by its SET domain, required for the HKMT activity. In undifferentiated myoblasts, endogenous Ezh2 was associated with the transcriptional regulator YY1. Both Ezh2 and YY1 were detected, with the deacetylase HDAC1, at genomic regions of silent muscle-specific genes. Their presence correlated with methylation of K27 of histone H3. YY1 was required for Ezh2 binding because RNA interference of YY1 abrogated chromatin recruitment of Ezh2 and prevented H3-K27 methylation. Upon gene activation, Ezh2, HDAC1, and YY1 dissociated from muscle loci, H3-K27 became hypomethylated and MyoD and SRF were recruited to the chromatin. These findings suggest the existence of a two-step activation mechanism whereby removal of H3-K27 methylation, conferred by an active Ezh2-containing protein complex, followed by recruitment of positive transcriptional regulators at discrete genomic loci are required to promote muscle gene expression and cell differentiation.
Figures
References
-
- Bannister A.J., Schneider, R., and Kouzarides, T. 2002. Histone methylation: Dynamic or static? Cell 109 801-806. - PubMed
-
- Bergstrom D.A., Penn, B.H., Strand, A., Perry, R.L., Rudnicki, M.A., and Tapscott, S.J. 2002. Promoter-specific regulation of MyoD binding and signal transduction cooperate to pattern gene expression. Mol. Cell 9 587-600. - PubMed
-
- Brown J.L., Mucci, D., Whiteley, M., Dirksen, M.L., and Kassis, J.A. 1998. The Drosophila Polycomb group gene pleiohomeotic encodes a DNA binding protein with homology to the transcription factor YY1. Mol. Cell 1 1057-1064. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous