

Serial segmental duplications during primate evolution result in complex human genome architecture
- PMID: 15520286
- PMCID: PMC525679
- DOI: 10.1101/gr.2746604
Serial segmental duplications during primate evolution result in complex human genome architecture
Abstract
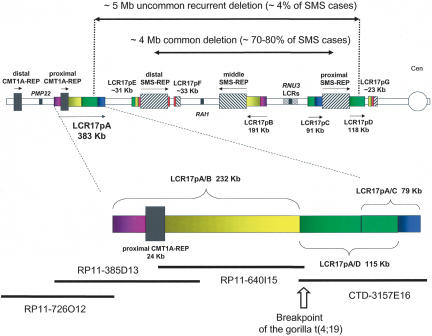
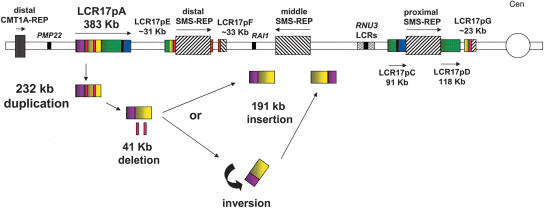
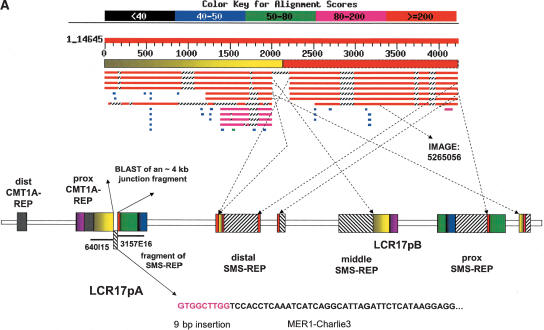
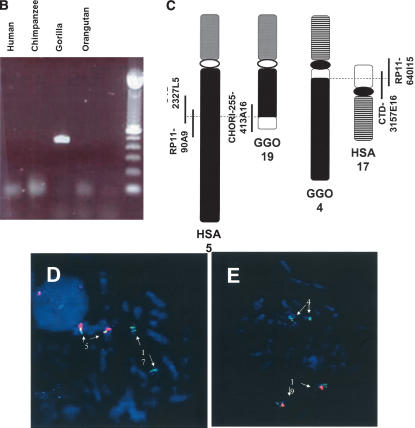
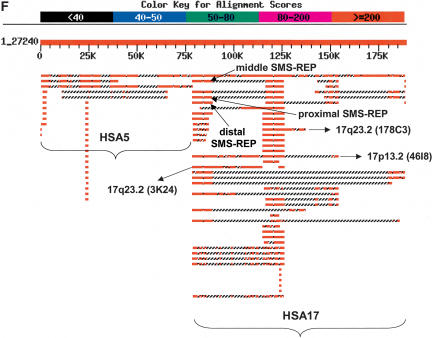
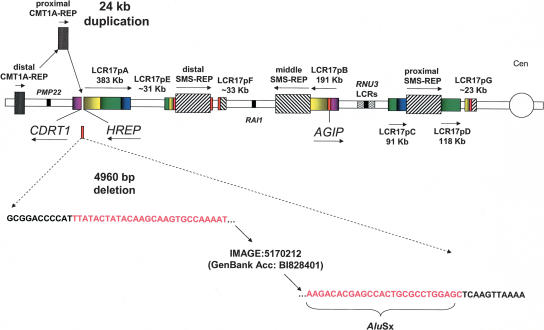
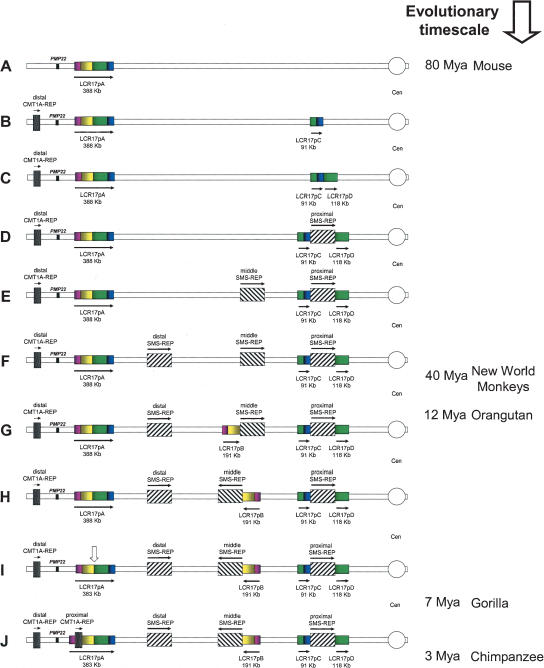
The human genome is particularly rich in low-copy repeats (LCRs) or segmental duplications (5%-10%), and this characteristic likely distinguishes us from lower mammals such as rodents. How and why the complex human genome architecture consisting of multiple LCRs has evolved remains an open question. Using molecular and computational analyses of human and primate genomic regions, we analyzed the structure and evolution of LCRs that resulted in complex architectural features of the human genome in proximal 17p. We found that multiple LCRs of different origins are situated adjacent to one another, whereas each LCR changed at different time points between >25 to 3-7 million years ago (Mya) during primate evolution. Evolutionary studies in primates suggested communication between the LCRs by gene conversion. The DNA transposable element MER1-Charlie3 and retroviral ERVL elements were identified at the breakpoint of the t(4;19) chromosome translocation in Gorilla gorilla, suggesting a potential role for transpositions in evolution of the primate genome. Thus, a series of consecutive segmental duplication events during primate evolution resulted in complex genome architecture in proximal 17p. Some of the more recent events led to the formation of novel genes that in human are expressed primarily in the brain. Our observations support the contention that serial segmental duplication events might have orchestrated primate evolution by the generation of novel fusion/fission genes as well as potentially by genomic inversions associated with decreased recombination rates facilitating gene divergence.
Figures
References
-
- Armengol, L., Pujana, M.A., Cheung, J., Scherer, S.W., and Estivill, X. 2003. Enrichment of segmental duplications in regions of breaks of synteny between the human and mouse genomes suggest their involvement in evolutionary rearrangements. Hum. Mol. Genet. 12: 2201-2208. - PubMed
WEB SITE REFERENCES
-
- www.chori.org/bacpac; Children's Hospital Oakland—BAC-PAC Resources.
-
- http://www.ncbi.nlm.nih.gov/; National Center for Biotechnology Information (NCBI).
-
- http://genome.ucsc.edu/; The UCSC genome bioinformatics site.
Publication types
MeSH terms
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources