

Essential role of membrane-attack protein in malarial transmission to mosquito host
- PMID: 15520375
- PMCID: PMC524694
- DOI: 10.1073/pnas.0406187101
Essential role of membrane-attack protein in malarial transmission to mosquito host
Abstract
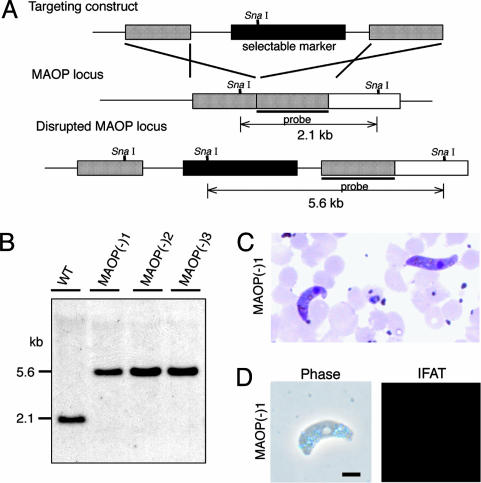
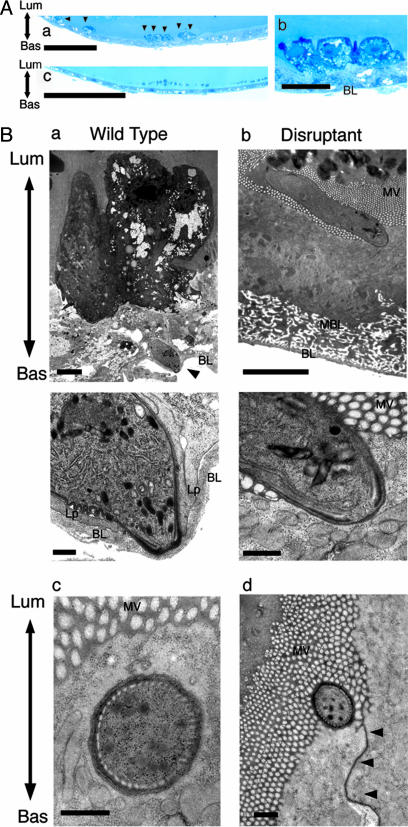
After ingestion of infected blood by a mosquito, malarial parasites are fertilized in the mosquito midgut and develop into motile ookinetes. These ookinetes invade epithelial cells by rupturing the cell membrane and migrate through the cytoplasm toward the basal lamina, on which they develop to oocysts. Here we report that a microneme protein with a membrane-attack complex and perforin (MACPF)-related domain, which we name membrane-attack ookinete protein (MAOP), is produced in the ookinete stage and plays an essential role in midgut invasion by the ookinete. Ookinetes with the MAOP gene disrupted completely lost infectivity to the midgut. After ingestion of blood infected with the disrupted parasite, the midgut epithelium remained intact, making a clear contrast with the damaged midgut epithelium invaded by wild-type ookinetes. Electron microscopic analysis showed that the disruptant ookinetes migrate to the gut epithelium and attach to the cell surface at the apical tip, but are unable to enter the cytoplasm by rupturing the cell membrane. These results indicate that the MAOP molecule acts on the plasma membrane of the host-cell-like mammalian MACPF family proteins that create pores in the membrane of target cells. Another previously identified MACPF-related molecule is produced in the liver-infective sporozoite and has a crucial role in traversing the liver sinusoidal cell boundary. The present finding, thus, suggests that conserved mechanisms for membrane rupture involving MACPF-related proteins are used in different host invasive stages of the malarial parasite, playing a key role in breaching biological barriers of host organs.
Figures

References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
