

Dopamine transporters depolarize neurons by a channel mechanism
- PMID: 15520385
- PMCID: PMC528740
- DOI: 10.1073/pnas.0403299101
Dopamine transporters depolarize neurons by a channel mechanism
Abstract
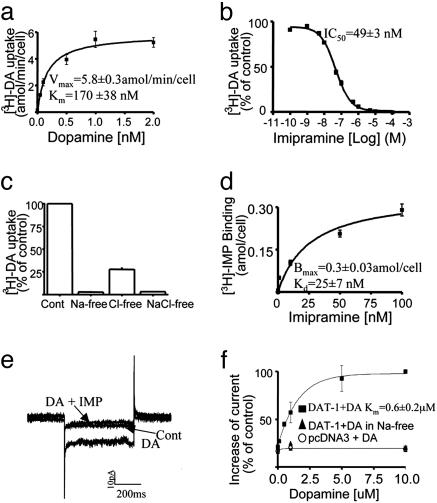
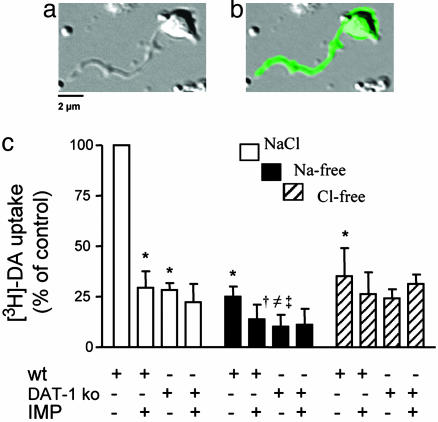
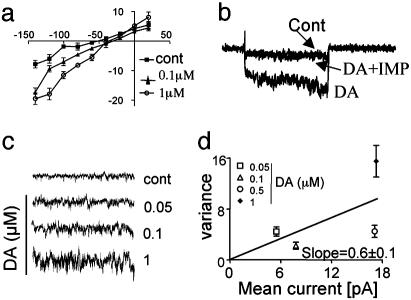
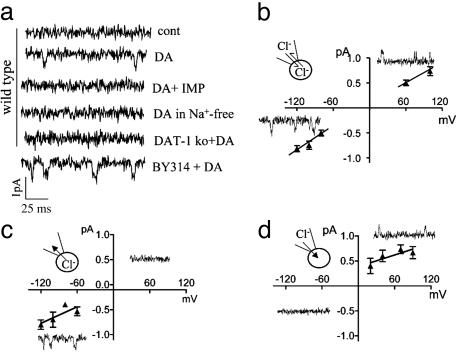
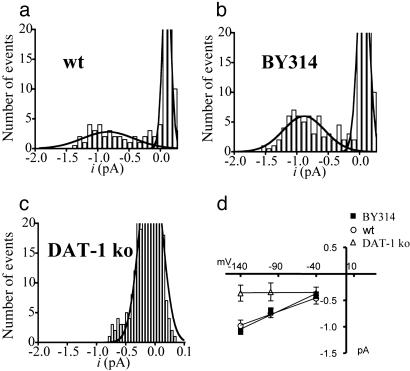
Neurotransmitter transporters generate larger currents than expected if one assumes fixed stoichiometry models. It remains controversial, however, whether these depolarizing currents arise from high density and rapid turnover rates of a classical transporter, or whether transporters exhibit bona fide channel behavior. Although heterologously expressed transporters show single-channel behavior and noise analysis in native cells strongly suggests channel behavior, no directly observed single-channel events associated with transporters have been reported thus far in native cells. We describe single-channel events arising directly from the Caenorhabditis elegans dopamine transporter (DAT-1) as evidenced by DA-induced channel activity blocked by a high-affinity DAT-1 inhibitor, increased channel activity in neurons that overexpress DAT-1, and loss of channels in dat-1 knockout neurons. Our data indicate that authentic transporter channels underlie depolarizing whole-cell currents. Thus, DA transporters not only transport DA but also exhibit a channel mode of conduction that directly modulates membrane potential and neuronal function.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
