

Cancer siRNA therapy by tumor selective delivery with ligand-targeted sterically stabilized nanoparticle
- PMID: 15520458
- PMCID: PMC528817
- DOI: 10.1093/nar/gnh140
Cancer siRNA therapy by tumor selective delivery with ligand-targeted sterically stabilized nanoparticle
Abstract
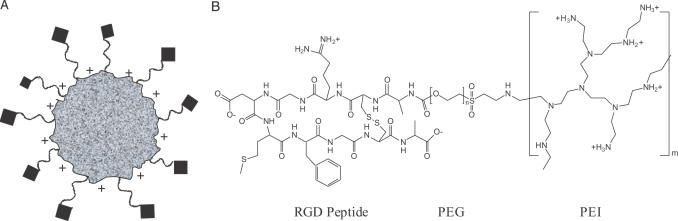
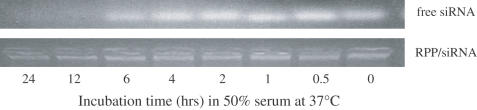
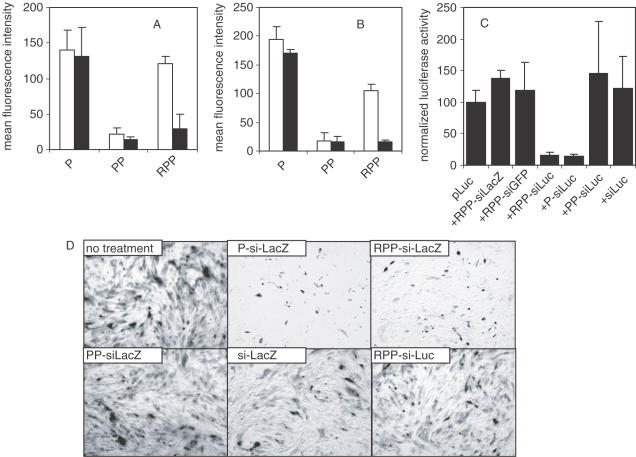
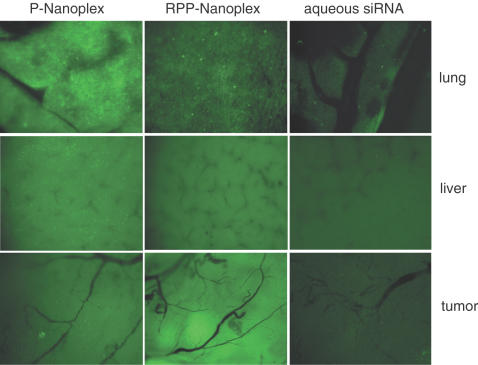
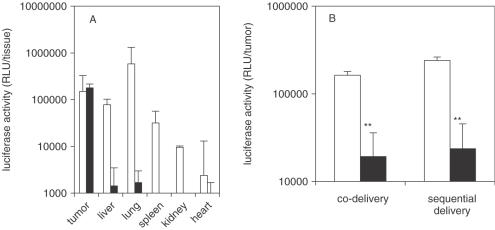
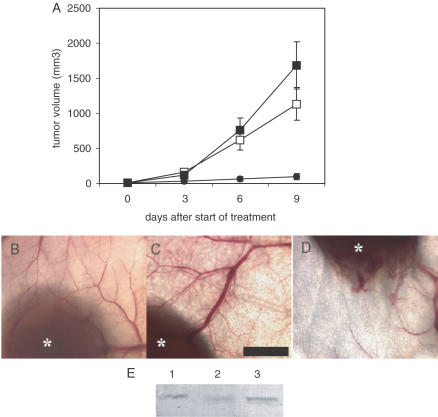
Potent sequence selective gene inhibition by siRNA 'targeted' therapeutics promises the ultimate level of specificity, but siRNA therapeutics is hindered by poor intracellular uptake, limited blood stability and non-specific immune stimulation. To address these problems, ligand-targeted, sterically stabilized nanoparticles have been adapted for siRNA. Self-assembling nanoparticles with siRNA were constructed with polyethyleneimine (PEI) that is PEGylated with an Arg-Gly-Asp (RGD) peptide ligand attached at the distal end of the polyethylene glycol (PEG), as a means to target tumor neovasculature expressing integrins and used to deliver siRNA inhibiting vascular endothelial growth factor receptor-2 (VEGF R2) expression and thereby tumor angiogenesis. Cell delivery and activity of PEGylated PEI was found to be siRNA sequence specific and depend on the presence of peptide ligand and could be competed by free peptide. Intravenous administration into tumor-bearing mice gave selective tumor uptake, siRNA sequence-specific inhibition of protein expression within the tumor and inhibition of both tumor angiogenesis and growth rate. The results suggest achievement of two levels of targeting: tumor tissue selective delivery via the nanoparticle ligand and gene pathway selectivity via the siRNA oligonucleotide. This opens the door for better targeted therapeutics with both tissue and gene selectivity, also to improve targeted therapies with less than ideal therapeutic targets.
Figures
References
-
- Ferrara N., Hillan,K.J., Gerber,H.-P. and Novotny,W. (2004) Discovery and development of bevacizumab, an anti-VEGF antibody for treating cancer. Nature Rev. Drug Discov., 3, 391–400. - PubMed
-
- Elbashir S.M., Harborth,J., Weber,K. and Tuschl,T. (2002) Analysis of gene function in somatic mammalian cells using small interfering RNAs. Methods, 26, 199–213. - PubMed
-
- Paddison P.J. and Hannon,G.J. (2003) siRNAs and shRNAs: skeleton keys to the human genome. Curr. Opin. Mol. Ther., 5, 217–224. - PubMed
-
- Carpenter A.E. and Sabatini,D.M. (2004) Systematic genome-wide screens of gene function. Nature Rev. Genet., 5, 11–22. - PubMed
-
- Ganju P. and Hall,J. (2004) Potential applications of siRNA for pain therapy. Expert Opin. Biol. Ther., 4, 531–542. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
