

Gain and loss of fruit flavor compounds produced by wild and cultivated strawberry species
- PMID: 15522848
- PMCID: PMC527202
- DOI: 10.1105/tpc.104.023895
Gain and loss of fruit flavor compounds produced by wild and cultivated strawberry species
Abstract
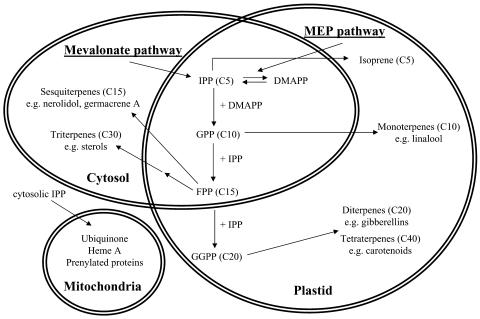
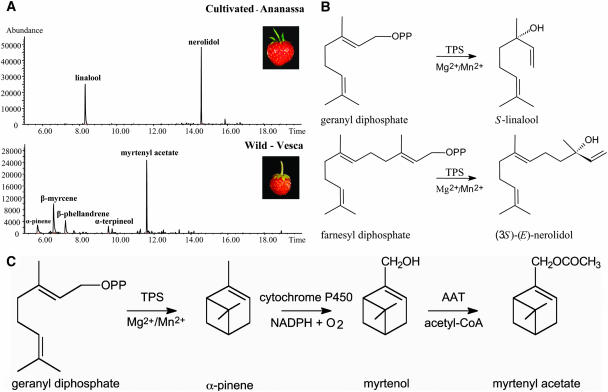
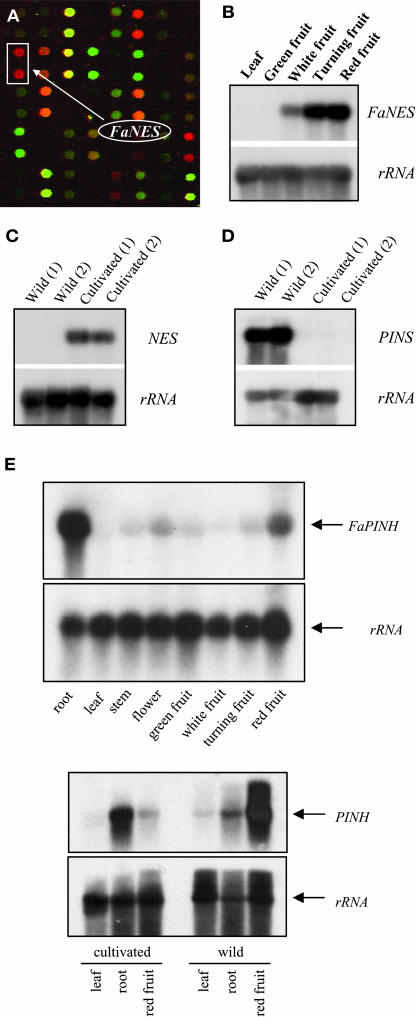
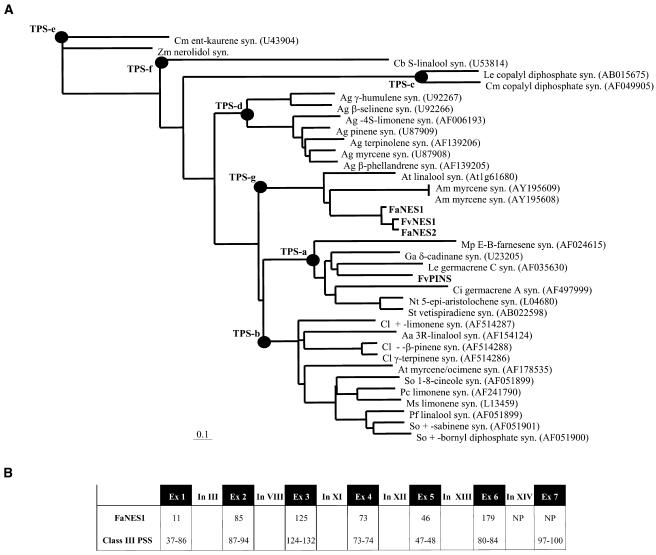
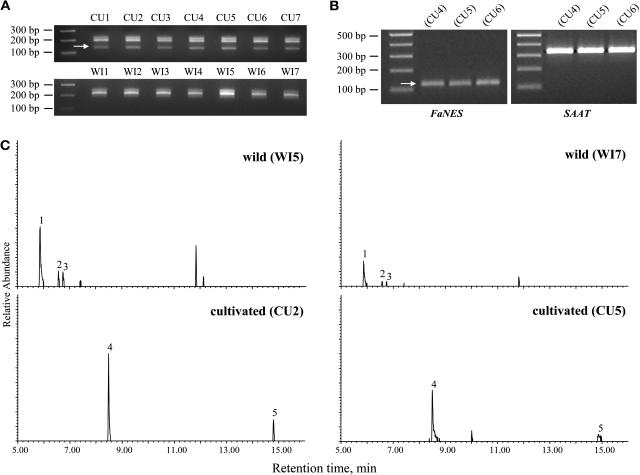
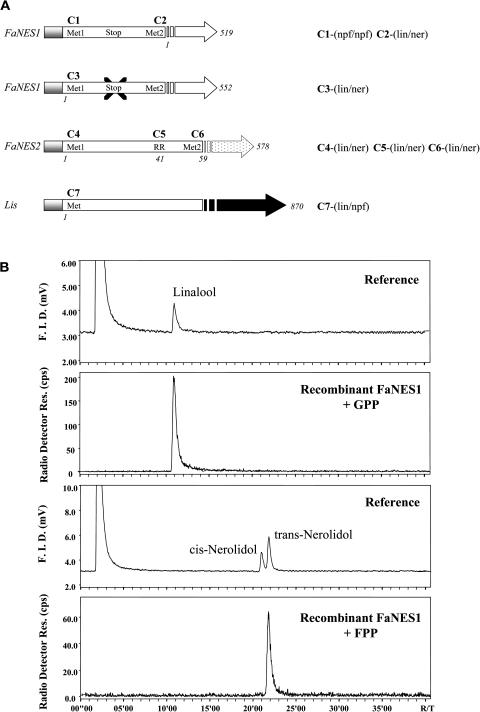
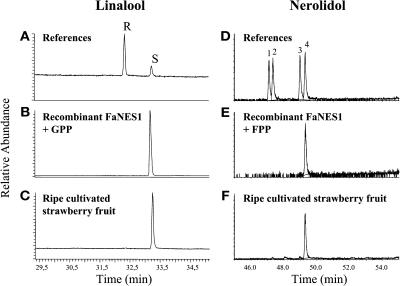
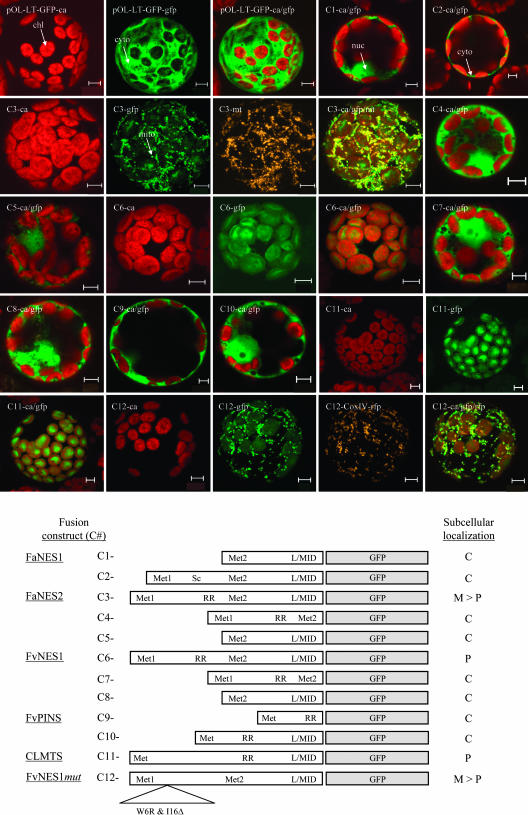
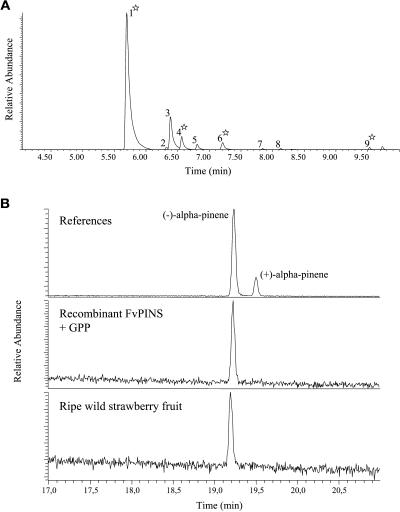
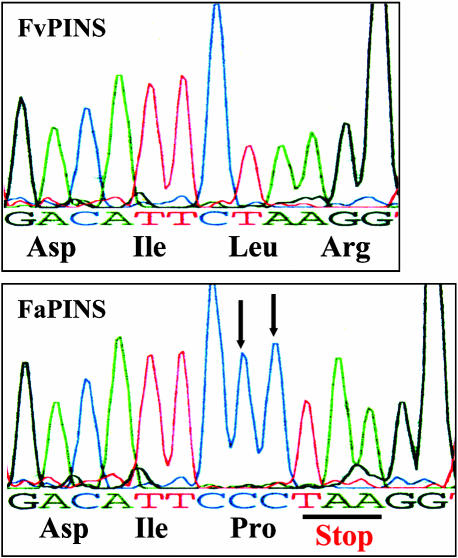
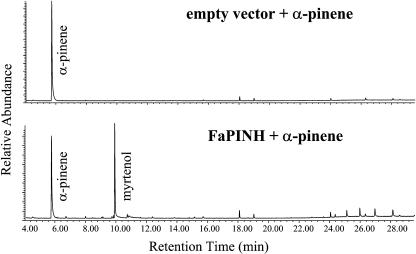
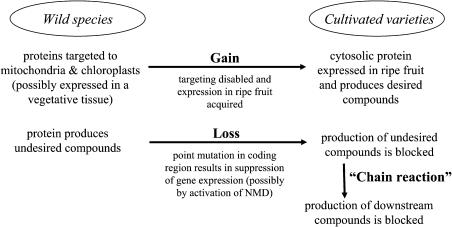
The blends of flavor compounds produced by fruits serve as biological perfumes used to attract living creatures, including humans. They include hundreds of metabolites and vary in their characteristic fruit flavor composition. The molecular mechanisms by which fruit flavor and aroma compounds are gained and lost during evolution and domestication are largely unknown. Here, we report on processes that may have been responsible for the evolution of diversity in strawberry (Fragaria spp) fruit flavor components. Whereas the terpenoid profile of cultivated strawberry species is dominated by the monoterpene linalool and the sesquiterpene nerolidol, fruit of wild strawberry species emit mainly olefinic monoterpenes and myrtenyl acetate, which are not found in the cultivated species. We used cDNA microarray analysis to identify the F. ananassa Nerolidol Synthase1 (FaNES1) gene in cultivated strawberry and showed that the recombinant FaNES1 enzyme produced in Escherichia coli cells is capable of generating both linalool and nerolidol when supplied with geranyl diphosphate (GPP) or farnesyl diphosphate (FPP), respectively. Characterization of additional genes that are very similar to FaNES1 from both the wild and cultivated strawberry species (FaNES2 and F. vesca NES1) showed that only FaNES1 is exclusively present and highly expressed in the fruit of cultivated (octaploid) varieties. It encodes a protein truncated at its N terminus. Green fluorescent protein localization experiments suggest that a change in subcellular localization led to the FaNES1 enzyme encountering both GPP and FPP, allowing it to produce linalool and nerolidol. Conversely, an insertional mutation affected the expression of a terpene synthase gene that differs from that in the cultivated species (termed F. ananassa Pinene Synthase). It encodes an enzyme capable of catalyzing the biosynthesis of the typical wild species monoterpenes, such as alpha-pinene and beta-myrcene, and caused the loss of these compounds in the cultivated strawberries. The loss of alpha-pinene also further influenced the fruit flavor profile because it was no longer available as a substrate for the production of the downstream compounds myrtenol and myrtenyl acetate. This phenomenon was demonstrated by cloning and characterizing a cytochrome P450 gene (Pinene Hydroxylase) that encodes the enzyme catalyzing the C10 hydroxylation of alpha-pinene to myrtenol. The findings shed light on the molecular evolutionary mechanisms resulting in different flavor profiles that are eventually selected for in domesticated species.
Figures


References
-
- Aharoni, A., and O'Connell, A.P. (2002). Gene expression analysis of strawberry achene and receptacle maturation using DNA microarrays. J. Exp. Bot. 53, 2073–2087. - PubMed
-
- Aubourg, S., Lecharny, A., and Bohlmann, J. (2002). Genomic analysis of the terpenoid synthase (AtTPS) gene family of Arabidopsis thaliana. Mol. Genet. Genomics 267, 730–745. - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
