

Layer-specific pyramidal cell oscillations evoked by tetanic stimulation in the rat hippocampal area CA1 in vitro and in vivo
- PMID: 15528242
- PMCID: PMC1665487
- DOI: 10.1113/jphysiol.2004.075390
Layer-specific pyramidal cell oscillations evoked by tetanic stimulation in the rat hippocampal area CA1 in vitro and in vivo
Abstract
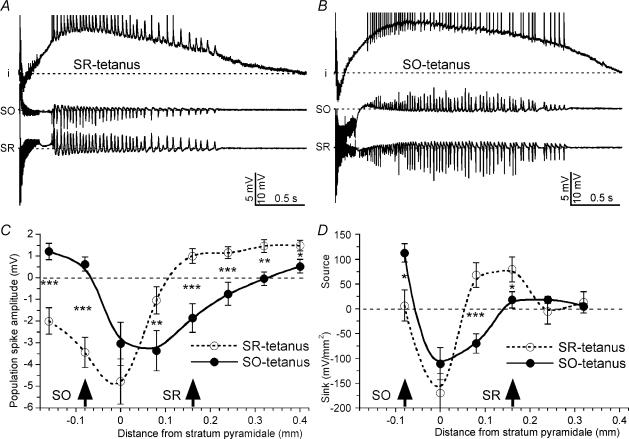
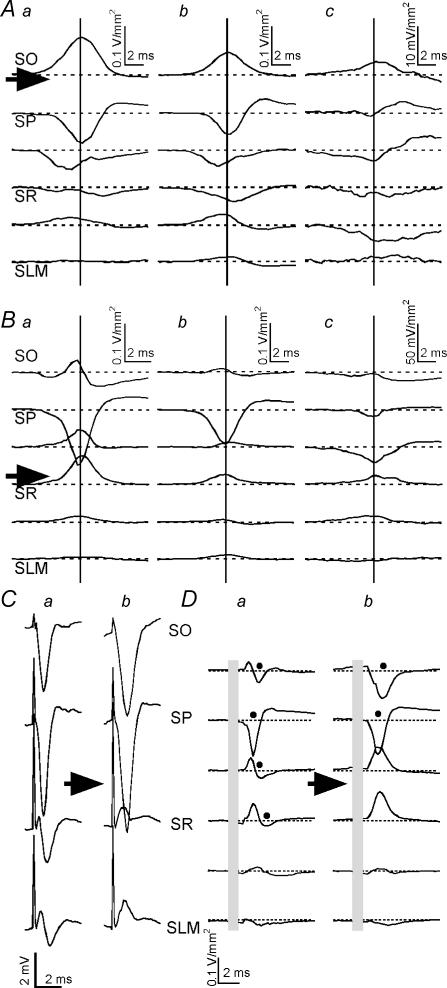
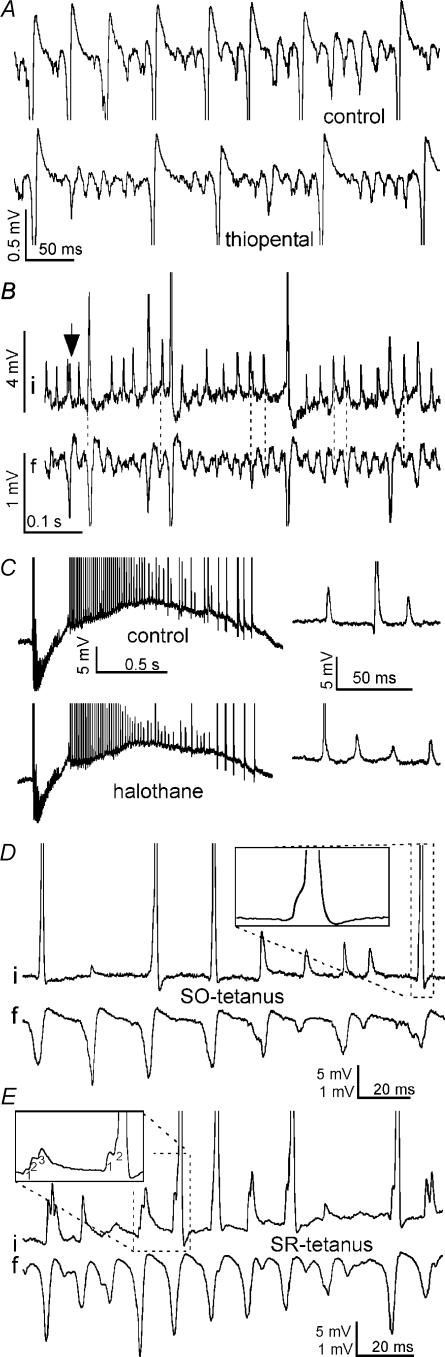
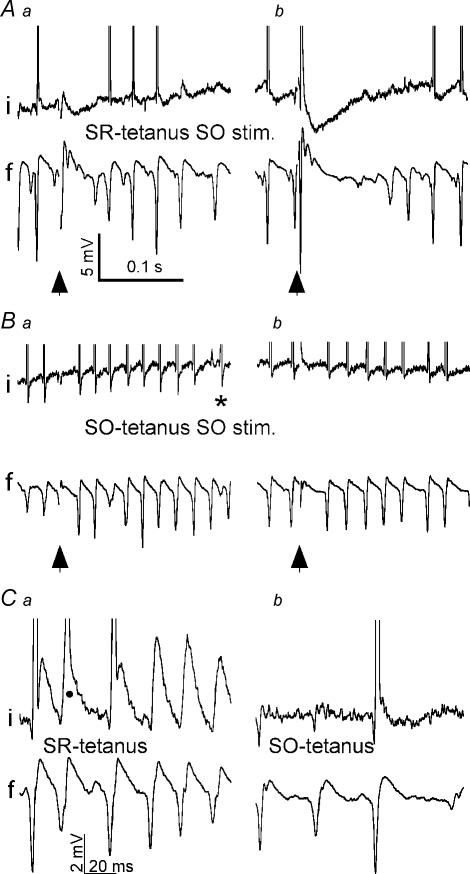
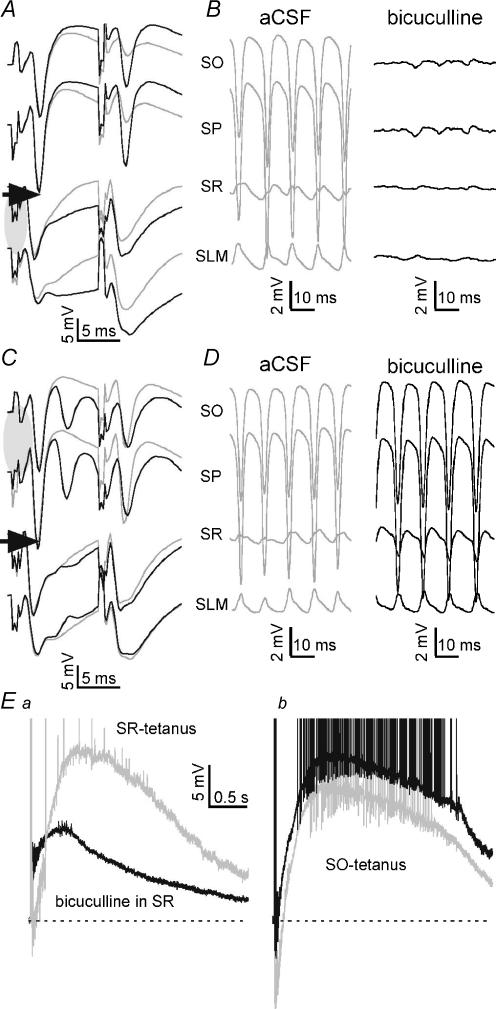
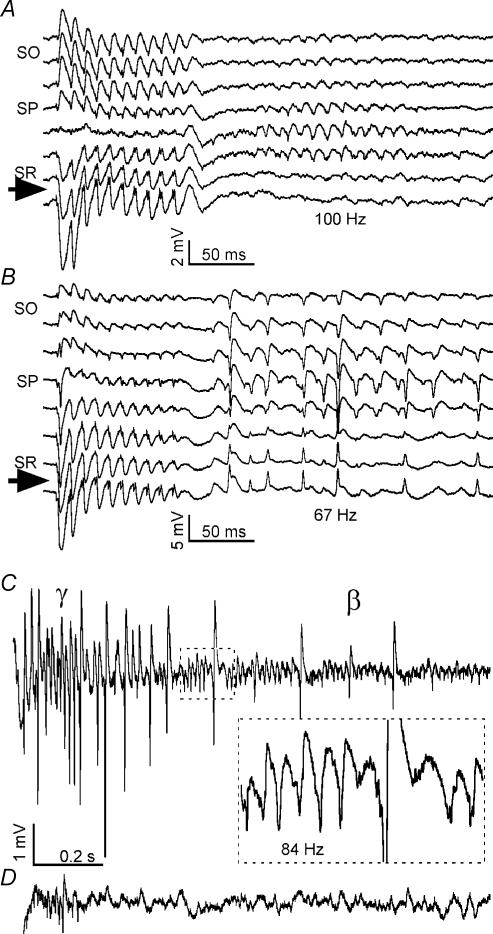
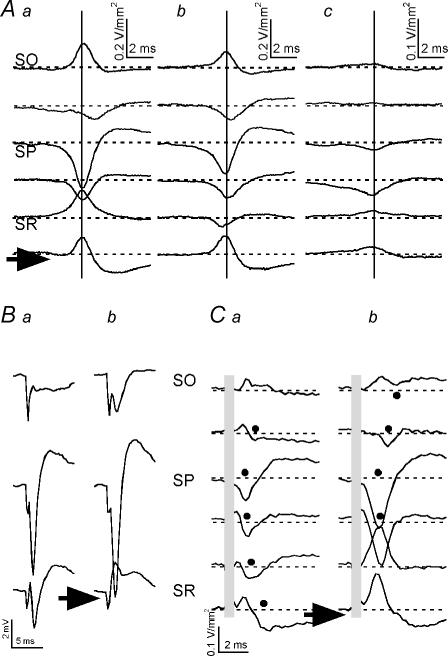
Tetanic stimulation of axons terminating in the CA1 region of the hippocampus induces oscillations in the gamma-to-beta frequency band (13-100 Hz) and can induce long-term potentiation (LTP). The rapid pyramidal cell discharge is driven by a mainly GABA(A)-receptor-mediated slow depolarization and entrained mainly through ephaptic interactions. This study tests whether cellular compartmentalization can explain how cells, despite severely reduced input resistance, can still fire briskly and have IPSPs superimposed on the slow GABAergic depolarization, and whether this behaviour occurs in vivo. Oscillations induced in CA1 in vitro by tetanic stimulation of the stratum radiatum or oriens were analysed using intracellular and multichannel field potentials along the cell axis. Layer-specific effects of focal application of bicuculline indicate that the GABAergic depolarization is concentrated on tetanized dendrites. Current-source density analysis and characteristics of partial spikes indicate that early action potentials are initiated in the proximal nontetanized dendrite but cannot invade the tetanized dendrite, where recurrent EPSPs and evoked IPSPs were largely suppressed. As the oscillation progresses, IPSPs recover and slow the neuronal firing to beta frequencies, with a small subpopulation of neurons continuing to fire at gamma frequency. Carbonic anhydrase dependence, threshold intensity, frequency, field strength and spike initiation/propagation of tetanus-evoked oscillations in urethane-anaesthetized rats, validate our observations in vitro, and show that these mechanisms operate in healthy tissue. However, the disrupted electrophysiology of the tetanized dendrites will disable normal information processing, has implications for LTP induction and is likely to play a role in pathological synchronization as found during epileptic discharges.
Figures
Similar articles
-
Limbic gamma rhythms. II. Synaptic and intrinsic mechanisms underlying spike doublets in oscillating subicular neurons.J Neurophysiol. 1998 Jul;80(1):162-71. doi: 10.1152/jn.1998.80.1.162. J Neurophysiol. 1998. PMID: 9658038
-
Long-term change in synaptic transmission in CA3 circuits followed by spontaneous rhythmic activity in rat hippocampal slices.Neurosci Res. 2001 Aug;40(4):325-36. doi: 10.1016/s0168-0102(01)00244-9. Neurosci Res. 2001. PMID: 11463478
-
Long-term suppression of synaptic transmission by tetanization of a single pyramidal cell in the mouse hippocampus in vitro.J Physiol. 1999 Mar 15;515 ( Pt 3)(Pt 3):757-67. doi: 10.1111/j.1469-7793.1999.757ab.x. J Physiol. 1999. PMID: 10066902 Free PMC article.
-
Modulation of the induction of long-term potentiation in the hippocampus.Adv Exp Med Biol. 1990;268:377-86. doi: 10.1007/978-1-4684-5769-8_41. Adv Exp Med Biol. 1990. PMID: 1963746 Review.
-
Generation of theta and gamma rhythms in the hippocampus.Neurosci Biobehav Rev. 1998 Mar;22(2):275-90. doi: 10.1016/s0149-7634(97)00014-6. Neurosci Biobehav Rev. 1998. PMID: 9579318 Review.
Cited by
-
Chronic stimulation of cultured neuronal networks boosts low-frequency oscillatory activity at theta and gamma with spikes phase-locked to gamma frequencies.J Neural Eng. 2012 Apr;9(2):026015. doi: 10.1088/1741-2560/9/2/026015. Epub 2012 Feb 23. J Neural Eng. 2012. PMID: 22361724 Free PMC article.
-
GABAergic synchronization in the limbic system and its role in the generation of epileptiform activity.Prog Neurobiol. 2011 Oct;95(2):104-32. doi: 10.1016/j.pneurobio.2011.07.003. Epub 2011 Jul 23. Prog Neurobiol. 2011. PMID: 21802488 Free PMC article. Review.
-
Field effects in the CNS play functional roles.Front Neural Circuits. 2010 May 18;4:15. doi: 10.3389/fncir.2010.00015. eCollection 2010. Front Neural Circuits. 2010. PMID: 20508749 Free PMC article.
-
Epileptiform activities in slices of hippocampus from mice after intra-hippocampal injection of kainic acid.J Physiol. 2008 Oct 15;586(20):4891-904. doi: 10.1113/jphysiol.2008.156281. Epub 2008 Aug 28. J Physiol. 2008. PMID: 18755752 Free PMC article.
-
Neurophysiological endophenotypes across bipolar and schizophrenia psychosis.Schizophr Bull. 2008 Jul;34(4):760-73. doi: 10.1093/schbul/sbn049. Epub 2008 May 22. Schizophr Bull. 2008. PMID: 18502737 Free PMC article. Review.
References
-
- Amaral DG. Emerging principles of intrinsic hippocampal organization. Curr Opin Neurobiol. 1993;3:225–229. 10.1016/0959-4388(93)90214-J. - DOI - PubMed
-
- Autere AM, Lamsa K, Kaila K, Taira T. Synaptic activation of GABAA receptors induces neuronal uptake of Ca2+ in adult rat hippocampal slices. J Neurophysiol. 1999;81:811–816. - PubMed
-
- Avoli M, Psarropoulou C, Tancredi V, Fueta Y. On the synchronous activity induced by 4-aminopyridine in the CA3 subfield of juvenile rat hippocampus. J Neurophysiol. 1993;70:1018–1029. - PubMed
-
- Bikson M, Hahn PJ, Fox JE, Jefferys JG. Depolarization block of neurons during maintenance of electrographic seizures. J Neurophysiol. 2003;90:2402–2408. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous