

Structural insights into the target specificity of plant invertase and pectin methylesterase inhibitory proteins
- PMID: 15528298
- PMCID: PMC535884
- DOI: 10.1105/tpc.104.025684
Structural insights into the target specificity of plant invertase and pectin methylesterase inhibitory proteins
Abstract
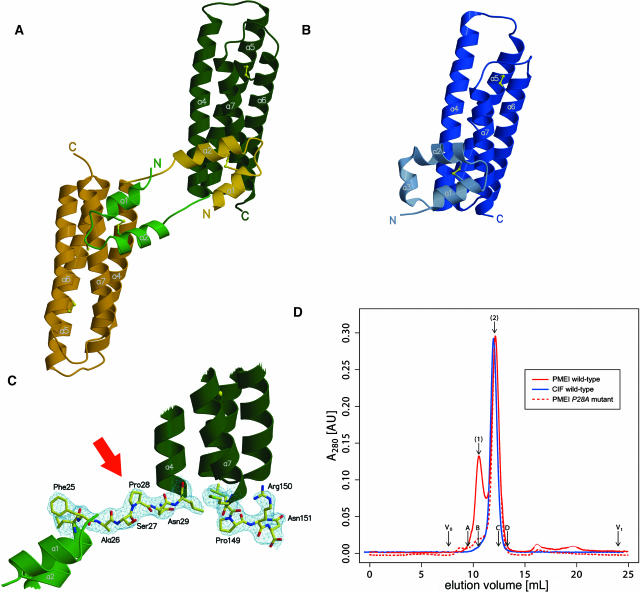
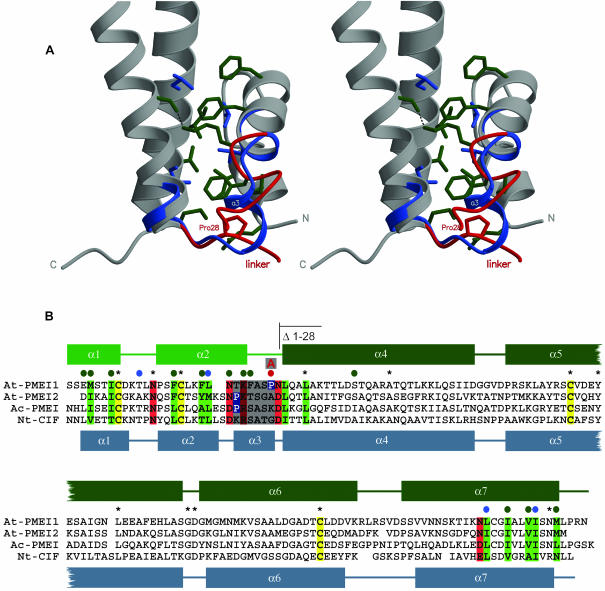
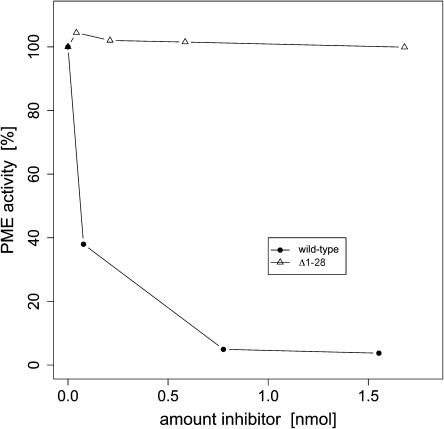
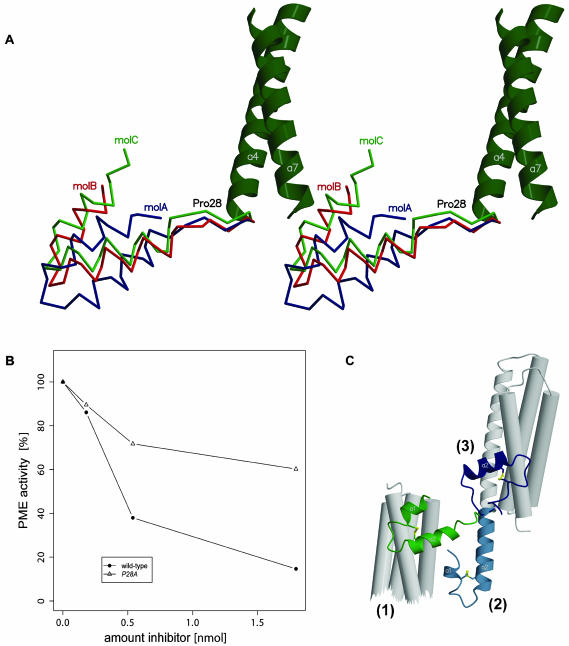
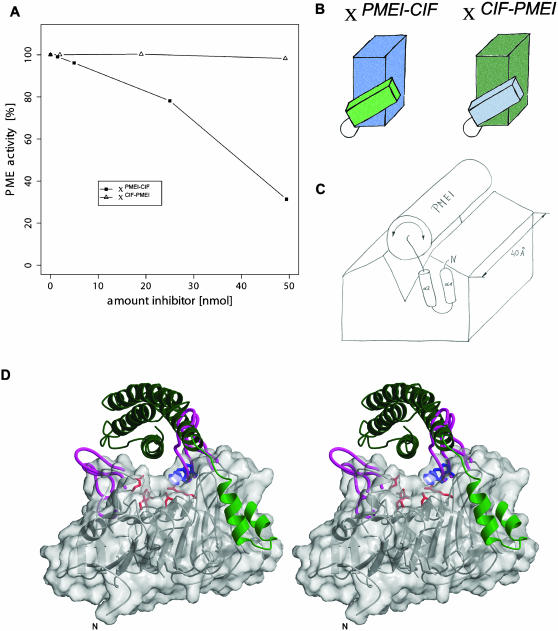
Pectin methylesterase (PME) and invertase are key enzymes in plant carbohydrate metabolism. Inhibitors of both enzymes constitute a sequence family of extracellular proteins. Members of this family are selectively targeted toward either PME or invertase. In a comparative structural approach we have studied how this target specificity is implemented on homologous sequences. By extending crystallographic work on the invertase inhibitor Nt-CIF to a pectin methylesterase inhibitor (PMEI) from Arabidopsis thaliana, we show an alpha-helical hairpin motif to be an independent and mobile structural entity in PMEI. Removal of this hairpin fully inactivates the inhibitor. A chimera composed of the alpha-hairpin of PMEI and the four-helix bundle of Nt-CIF is still active against PME. By contrast, combining the corresponding segment of Nt-CIF with the four-helix bundle of PMEI renders the protein inactive toward either PME or invertase. Our experiments provide insight in how these homologous inhibitors can make differential use of similar structural modules to achieve distinct functions. Integrating our results with previous findings, we present a model for the PME-PMEI complex with important implications.
Figures
Similar articles
-
Structural insights into the pH-controlled targeting of plant cell-wall invertase by a specific inhibitor protein.Proc Natl Acad Sci U S A. 2010 Oct 5;107(40):17427-32. doi: 10.1073/pnas.1004481107. Epub 2010 Sep 21. Proc Natl Acad Sci U S A. 2010. PMID: 20858733 Free PMC article.
-
Pectin methylesterase inhibitor.Biochim Biophys Acta. 2004 Feb 12;1696(2):245-52. doi: 10.1016/j.bbapap.2003.08.011. Biochim Biophys Acta. 2004. PMID: 14871665 Review.
-
The plant invertase inhibitor shares structural properties and disulfide bridges arrangement with the pectin methylesterase inhibitor.J Protein Chem. 2003 May;22(4):363-9. doi: 10.1023/a:1025342207831. J Protein Chem. 2003. PMID: 13678300
-
Activity of an atypical Arabidopsis thaliana pectin methylesterase.Planta. 2009 Jan;229(2):311-21. doi: 10.1007/s00425-008-0831-0. Epub 2008 Oct 21. Planta. 2009. PMID: 18936961
-
Plant protein inhibitors of invertases.Biochim Biophys Acta. 2004 Feb 12;1696(2):253-61. doi: 10.1016/j.bbapap.2003.09.017. Biochim Biophys Acta. 2004. PMID: 14871666 Review.
Cited by
-
External application of gametophyte-specific ZmPMEI1 induces pollen tube burst in maize.Plant Reprod. 2013 Sep;26(3):255-66. doi: 10.1007/s00497-013-0221-z. Epub 2013 Jul 4. Plant Reprod. 2013. PMID: 23824238
-
PpINH1, an invertase inhibitor, interacts with vacuolar invertase PpVIN2 in regulating the chilling tolerance of peach fruit.Hortic Res. 2020 Oct 1;7:168. doi: 10.1038/s41438-020-00389-8. eCollection 2020. Hortic Res. 2020. PMID: 33082974 Free PMC article.
-
The three-dimensional structure of the extracellular adhesion domain of the sialic acid-binding adhesin SabA from Helicobacter pylori.J Biol Chem. 2014 Mar 7;289(10):6332-6340. doi: 10.1074/jbc.M113.513135. Epub 2013 Dec 27. J Biol Chem. 2014. PMID: 24375407 Free PMC article.
-
Transcriptome sequencing and whole genome expression profiling of hexaploid sweetpotato under salt stress.BMC Genomics. 2020 Mar 4;21(1):197. doi: 10.1186/s12864-020-6524-1. BMC Genomics. 2020. PMID: 32131729 Free PMC article.
-
Cell wall/vacuolar inhibitor of fructosidase 1 regulates ABA response and salt tolerance in Arabidopsis.Plant Signal Behav. 2020 Apr 2;15(4):1744293. doi: 10.1080/15592324.2020.1744293. Epub 2020 Mar 26. Plant Signal Behav. 2020. PMID: 32213123 Free PMC article.
References
-
- Bode, W., and Huber, R. (1992). Natural protein proteinase inhibitors and their interaction with proteinases. Eur. J. Biochem. 204, 433–451. - PubMed
-
- Brummell, D.A., and Harpster, M.H. (2001). Cell wall metabolism in fruit softening and quality and its manipulation in transgenic plants. Plant Mol. Biol. 47, 311–340. - PubMed
-
- Brunger, A.T., et al. (1998). Crystallography & NMR system: A new software suite for macromolecular structure determination. Acta Crystallogr. D Biol. Crystallogr. 54, 905–921. - PubMed
-
- Camardella, L., Carratore, V., Ciardiello, M.A., Servillo, L., Balestrieri, C., and Giovane, A. (2000). Kiwi protein inhibitor of pectin methylesterase amino-acid sequence and structural importance of two disulfide bridges. Eur. J. Biochem. 267, 4561–4565. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
