

Human ribonuclease A superfamily members, eosinophil-derived neurotoxin and pancreatic ribonuclease, induce dendritic cell maturation and activation
- PMID: 15528350
- PMCID: PMC2847482
- DOI: 10.4049/jimmunol.173.10.6134
Human ribonuclease A superfamily members, eosinophil-derived neurotoxin and pancreatic ribonuclease, induce dendritic cell maturation and activation
Abstract
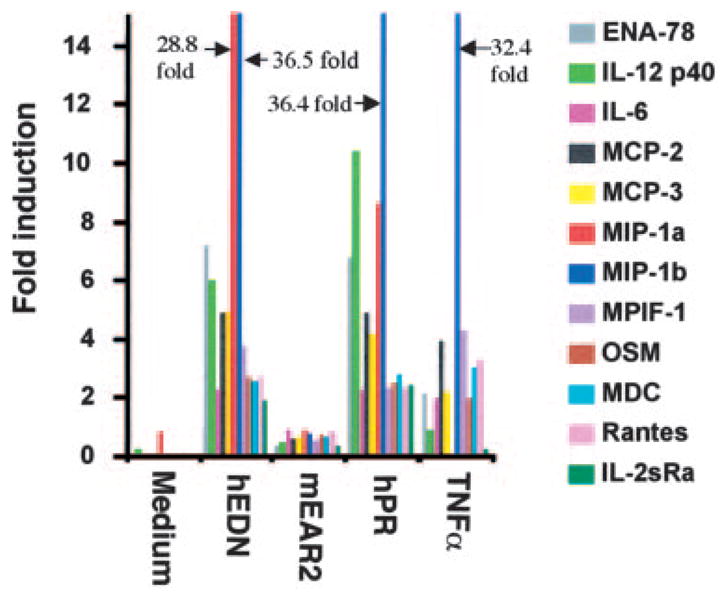
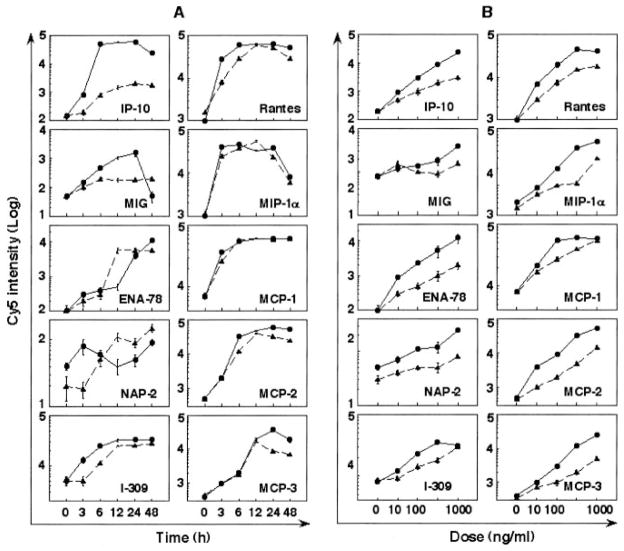
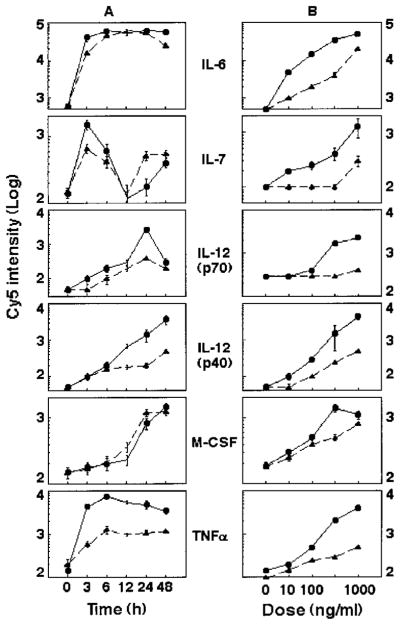
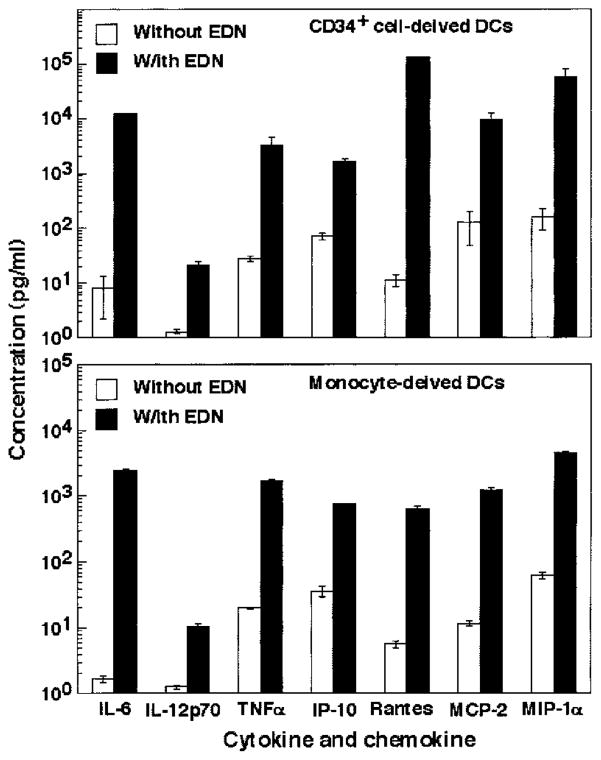
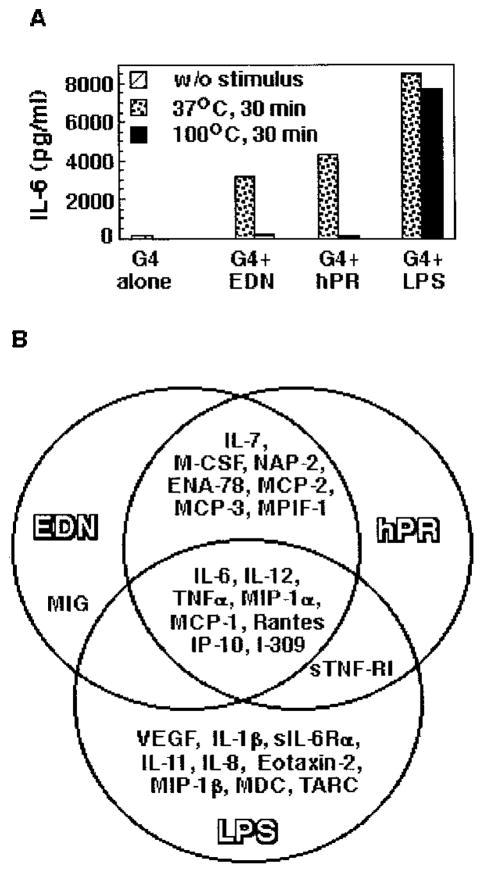
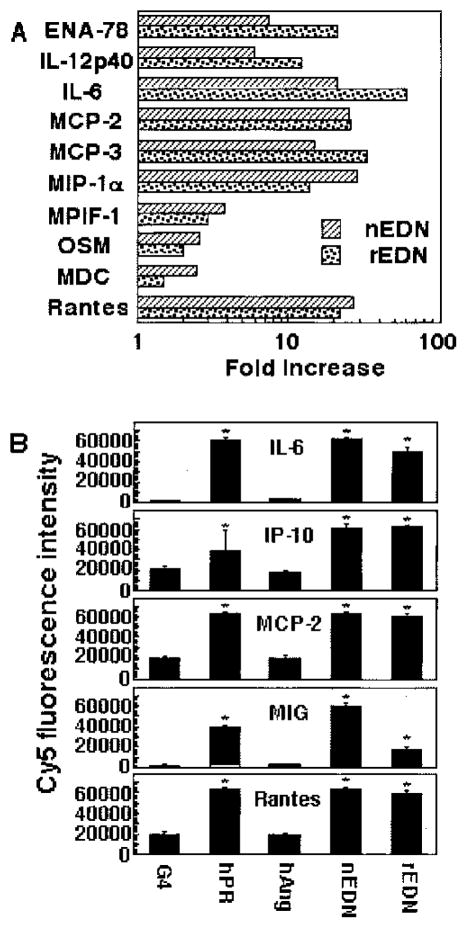
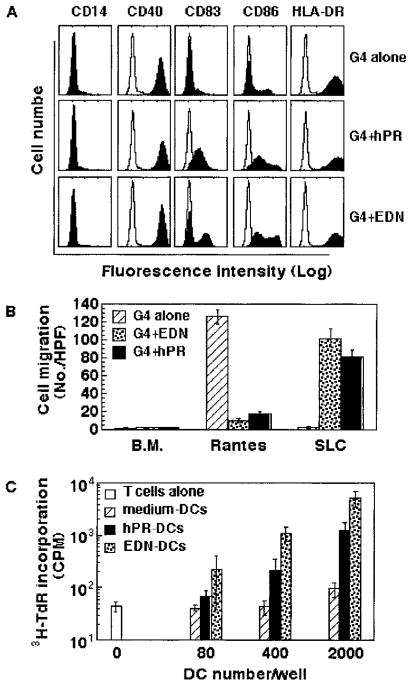
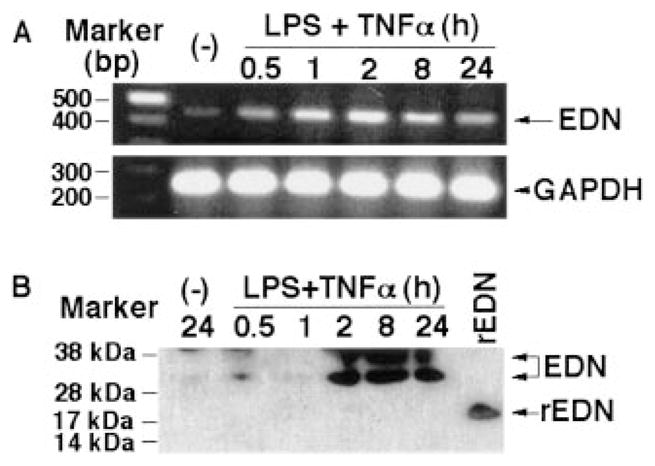
A number of mammalian antimicrobial proteins produced by neutrophils and cells of epithelial origin have chemotactic and activating effects on host cells, including cells of the immune system. Eosinophil granules contain an antimicrobial protein known as eosinophil-derived neurotoxin (EDN), which belongs to the RNase A superfamily. EDN has antiviral and chemotactic activities in vitro. In this study, we show that EDN, and to a lesser extent human pancreatic RNase (hPR), another RNase A superfamily member, activates human dendritic cells (DCs), leading to the production of a variety of inflammatory cytokines, chemokines, growth factors, and soluble receptors. Human angiogenin, a RNase evolutionarily more distant to EDN and hPR, did not display such activating effects. Additionally, EDN and hPR also induced phenotypic and functional maturation DCs. These RNases were as efficacious as TNF-alpha, but induced a different set of cytokine mediators. Furthermore, EDN production by human macrophages could be induced by proinflammatory stimuli. The results reveal the DC-activating activity of EDN and hPR and suggest that they are likely participants of inflammatory and immune responses. A number of endogenous mediators in addition to EDN have been reported to have both chemotactic and activating effects on APCs, and can thus amplify innate and Ag-specific immune responses to danger signals. We therefore propose these mediators be considered as endogenous multifunctional immune alarmins.
Figures
References
-
- Zasloff M. Antimicrobial peptides of multicellular organisms. Nature. 2002;415:389. - PubMed
-
- Scott MG, Hancock REW. Cationic antimicrobial peptides and their multifunctional role in the immune system. Crit Rev Immunol. 2002;20:407. - PubMed
-
- Chertov O, Michiel DF, Xu L, Wang JM, Tani K, Murphy WJ, Longo DL, Taub DD, Oppenheim JJ. Identification of defensin-1, defensin-2, and CAP37/azurocidin as T-cell chemoattractant proteins released from interleukin-8-stimulated neutrophils. J Biol Chem. 1996;271:2935. - PubMed
-
- Yang D, Chen Q, Chertov O, Oppenheim JJ. Human neutrophil defensins selectively chemoattract naive T and immature dendritic cells. J Leukocyte Biol. 2000;68:9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
