

Analysis of sequence, map position, and gene expression reveals conserved essential genes for iron uptake in Arabidopsis and tomato
- PMID: 15531708
- PMCID: PMC535847
- DOI: 10.1104/pp.104.047233
Analysis of sequence, map position, and gene expression reveals conserved essential genes for iron uptake in Arabidopsis and tomato
Abstract
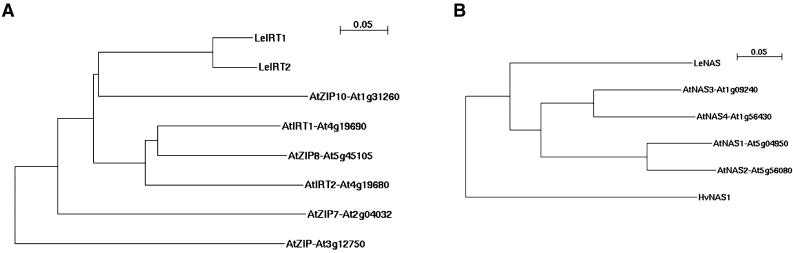
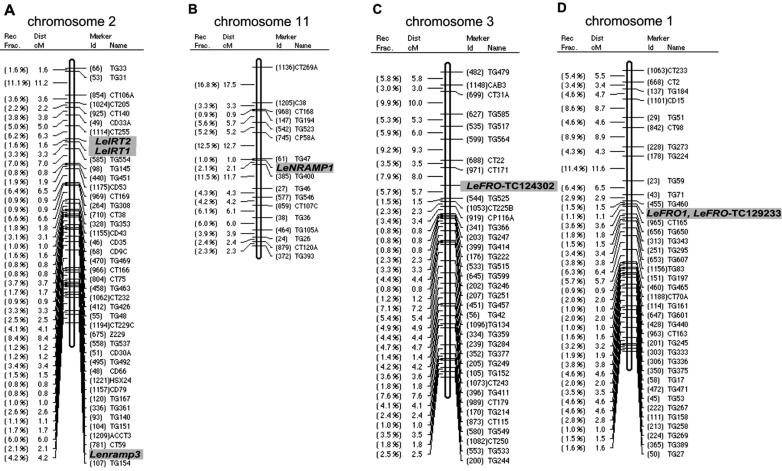
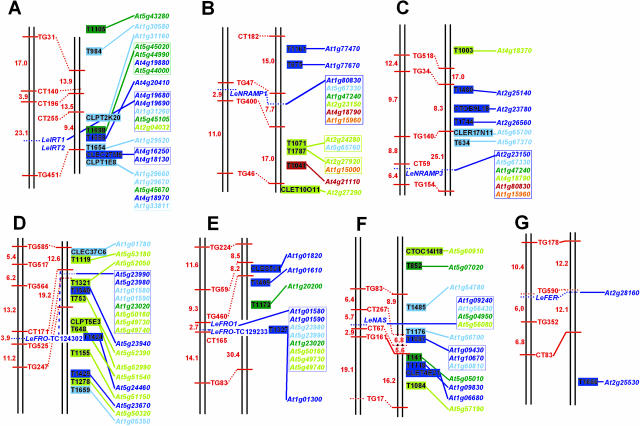
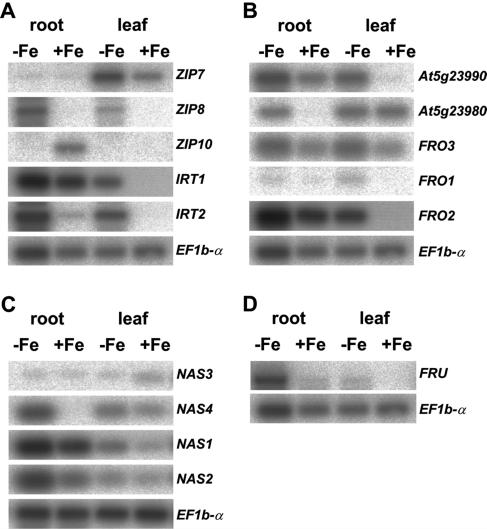
Arabidopsis (Arabidopsis thaliana) and tomato (Lycopersicon esculentum) show similar physiological responses to iron deficiency, suggesting that homologous genes are involved. Essential gene functions are generally considered to be carried out by orthologs that have remained conserved in sequence and map position in evolutionarily related species. This assumption has not yet been proven for plant genomes that underwent large genome rearrangements. We addressed this question in an attempt to deduce functional gene pairs for iron reduction, iron transport, and iron regulation between Arabidopsis and tomato. Iron uptake processes are essential for plant growth. We investigated iron uptake gene pairs from tomato and Arabidopsis, namely sequence, conserved gene content of the regions containing iron uptake homologs based on conserved orthologous set marker analysis, gene expression patterns, and, in two cases, genetic data. Compared to tomato, the Arabidopsis genome revealed more and larger gene families coding for the iron uptake functions. The number of possible homologous pairs was reduced if functional expression data were taken into account in addition to sequence and map position. We predict novel homologous as well as partially redundant functions of ferric reductase-like and iron-regulated transporter-like genes in Arabidopsis and tomato. Arabidopsis nicotianamine synthase genes encode a partially redundant family. In this study, Arabidopsis gene redundancy generally reflected the presumed genome duplication structure. In some cases, statistical analysis of conserved gene regions between tomato and Arabidopsis suggested a common evolutionary origin. Although involvement of conserved genes in iron uptake was found, these essential genes seem to be of paralogous rather than orthologous origin in tomato and Arabidopsis.
Figures

References
-
- Arabidopsis Genome Initiative (2000) Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408: 796–815 - PubMed
-
- Bauer P, Bereczky Z (2003) Gene networks involved in iron acquisition strategies in plants. Agronomie 23: 1–8
-
- Becher M, Talke IN, Krall L, Kramer U (2004) Cross-species microarray transcript profiling reveals high constitutive expression of metal homeostasis genes in shoots of the zinc hyperaccumulator Arabidopsis halleri. Plant J 37: 251–268 - PubMed
-
- Bereczky Z, Wang HY, Schubert V, Ganal M, Bauer P (2003) Differential regulation of nramp and irt metal transporter genes in wild type and iron uptake mutants of tomato. J Biol Chem 278: 24697–24704 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
