

Recycling endosomes can serve as intermediates during transport from the Golgi to the plasma membrane of MDCK cells
- PMID: 15534004
- PMCID: PMC2172492
- DOI: 10.1083/jcb.200408165
Recycling endosomes can serve as intermediates during transport from the Golgi to the plasma membrane of MDCK cells
Abstract
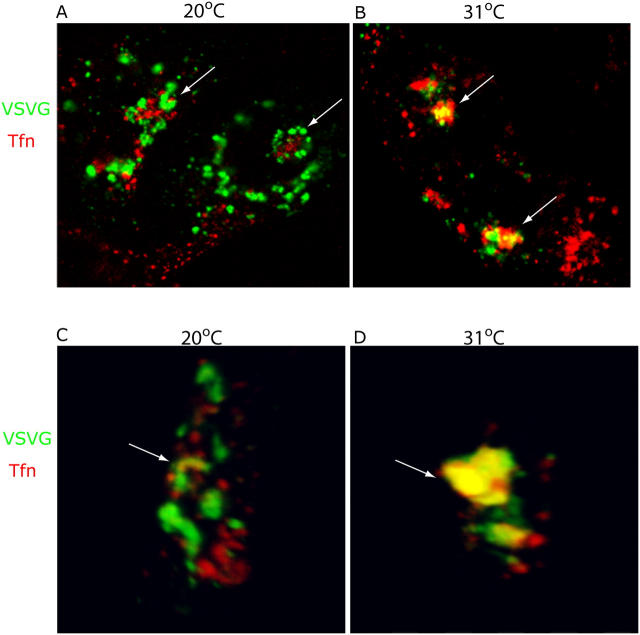
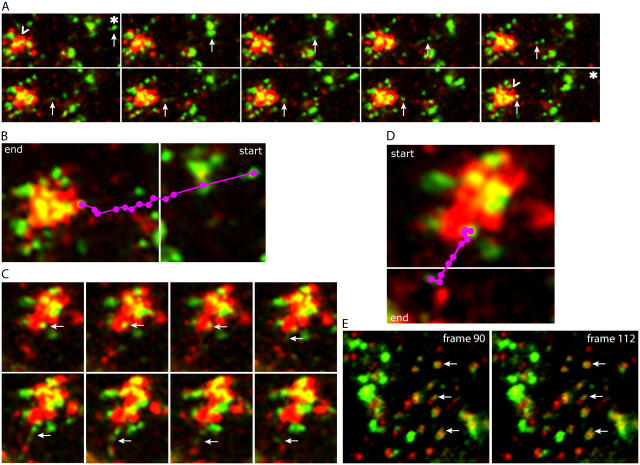
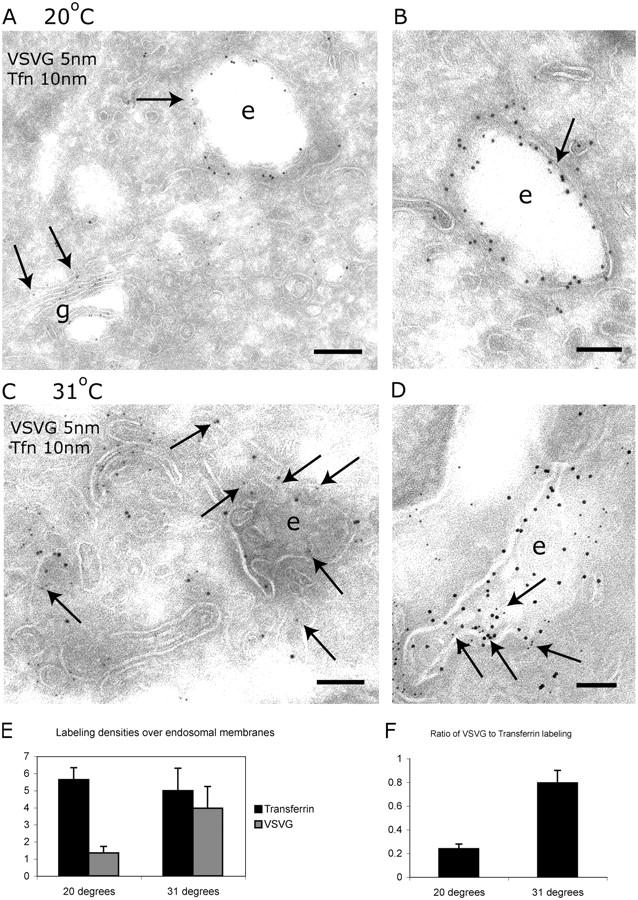
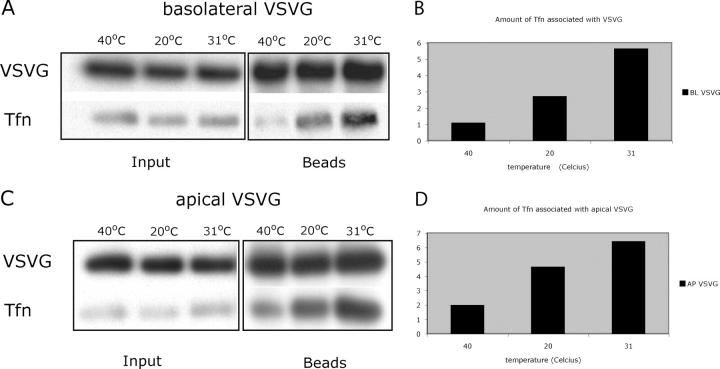
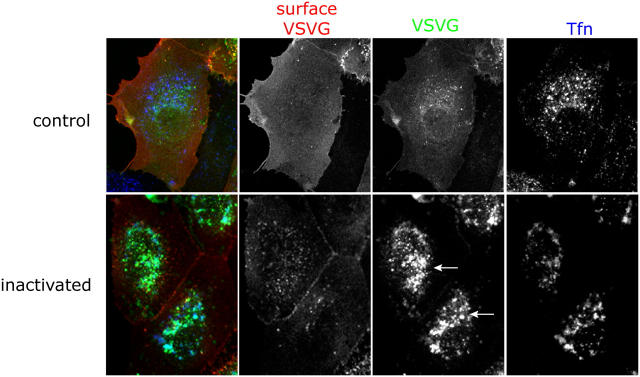
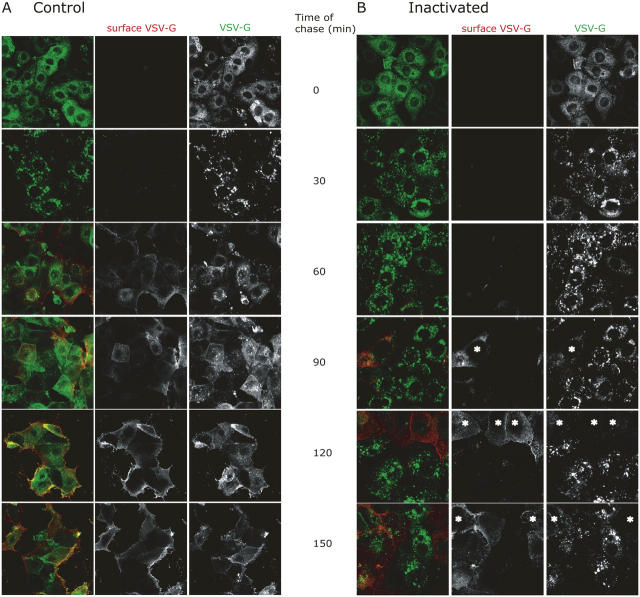
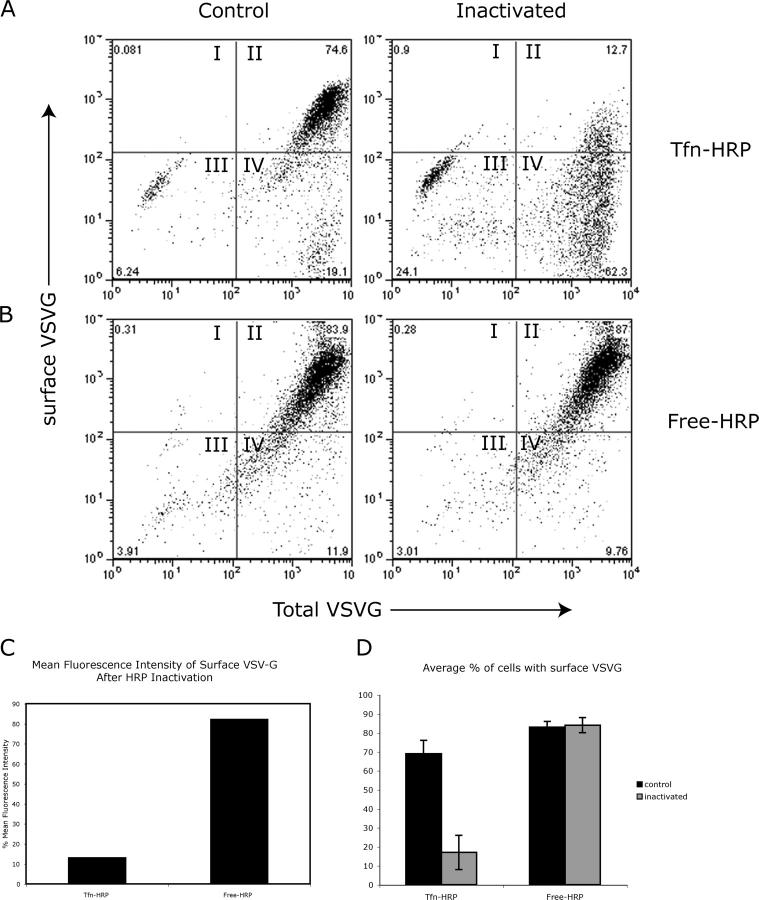
The AP-1B clathrin adaptor complex is responsible for the polarized transport of many basolateral membrane proteins in epithelial cells. Localization of AP-1B to recycling endosomes (REs) along with other components (exocyst subunits and Rab8) involved in AP-1B-dependent transport suggested that RE might be an intermediate between the Golgi and the plasma membrane. Although the involvement of endosomes in the secretory pathway has long been suspected, we now present direct evidence using four independent methods that REs play a role in basolateral transport in MDCK cells. Newly synthesized AP-1B-dependent cargo, vesicular stomatitis virus glycoprotein G (VSV-G), was found by video microscopy, immunoelectron microscopy, and cell fractionation to enter transferrin-positive REs within a few minutes after exit from the trans-Golgi network. Although transient, RE entry appears essential because enzymatic inactivation of REs blocked VSV-G delivery to the cell surface. Because an apically targeted VSV-G mutant behaved similarly, these results suggest that REs not only serve as an intermediate but also as a common site for polarized sorting on the endocytic and secretory pathways.
Figures
References
-
- Bonifacino, J.S., and L.M. Traub. 2003. Signals for sorting of transmembrane proteins to endosomes and lysosomes. Annu. Rev. Biochem. 72:395–447. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
