

A crucial arginine residue is required for a conformational switch in NifL to regulate nitrogen fixation in Azotobacter vinelandii
- PMID: 15534211
- PMCID: PMC528952
- DOI: 10.1073/pnas.0405312101
A crucial arginine residue is required for a conformational switch in NifL to regulate nitrogen fixation in Azotobacter vinelandii
Abstract
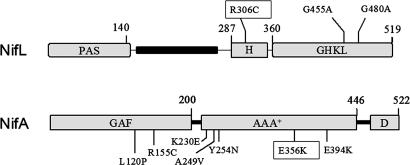
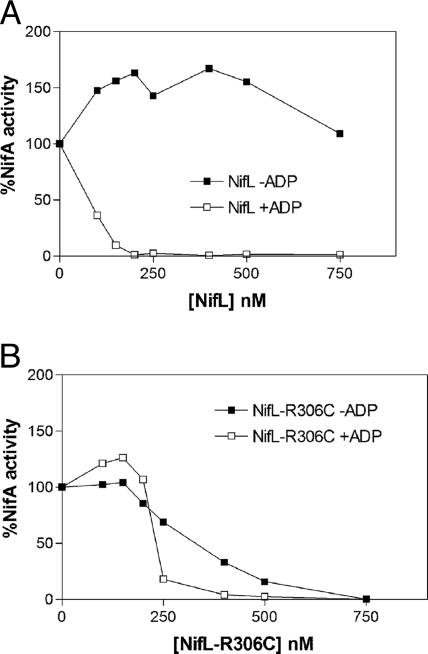
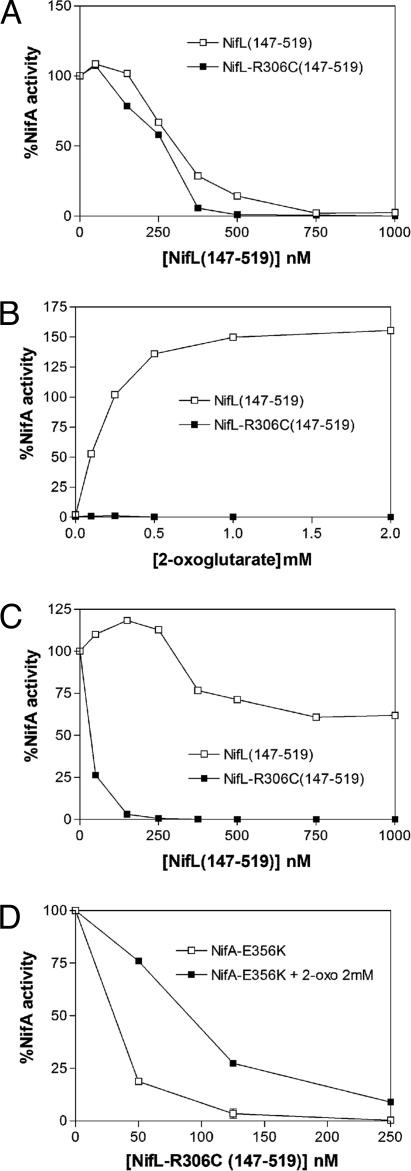
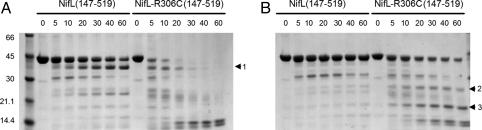
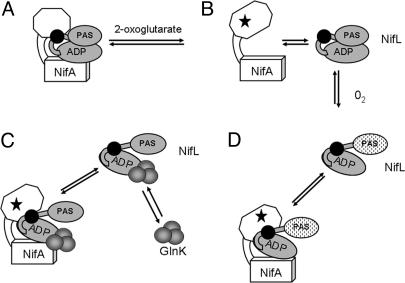
NifL is an antiactivator that tightly regulates transcription of genes required for nitrogen fixation in Azotobacter vinelandii by controlling the activity of its partner protein NifA, a member of the family of sigma(54)-dependent transcriptional activators. Although the C-terminal region of A. vinelandii NifL shows homology to the transmitter domains of histidine protein kinases, signal transduction between NifL and NifA is conveyed by means of protein-protein interactions rather than by phosphorylation. Binding of the ligand 2-oxoglutarate to NifA plays a crucial role in preventing inhibition by NifL under conditions appropriate for nitrogen fixation. We have used a suppressor screen to identify a critical arginine residue (R306) in NifL that is required to release NifA from inhibition under appropriate environmental conditions. Amino acid substitutions at position 306 result in constitutive inhibition of NifA activity by NifL, thus preventing nitrogen fixation. Biochemical studies with one of the mutant proteins demonstrate that the substitution alters the conformation of NifL significantly and prevents the response of NifA to 2-oxoglutarate. We propose that arginine 306 is critical for the propagation of signals perceived by A. vinelandii NifL in response to the redox and fixed-nitrogen status and is required for a conformational switch that inactivates the inhibitory function of NifL under conditions appropriate for nitrogen fixation.
Figures
Similar articles
-
Role of the amino-terminal GAF domain of the NifA activator in controlling the response to the antiactivator protein NifL.Mol Microbiol. 2004 Jun;52(6):1731-44. doi: 10.1111/j.1365-2958.2004.04089.x. Mol Microbiol. 2004. PMID: 15186421
-
Regulation of nitrogen fixation in Klebsiella pneumoniae and Azotobacter vinelandii: NifL, transducing two environmental signals to the nif transcriptional activator NifA.J Mol Microbiol Biotechnol. 2002 May;4(3):235-42. J Mol Microbiol Biotechnol. 2002. PMID: 11931553 Review.
-
Role of the H domain of the histidine kinase-like protein NifL in signal transmission.J Biol Chem. 2007 May 4;282(18):13429-37. doi: 10.1074/jbc.M610827200. Epub 2007 Mar 13. J Biol Chem. 2007. PMID: 17355964
-
Role of Escherichia coli nitrogen regulatory genes in the nitrogen response of the Azotobacter vinelandii NifL-NifA complex.J Bacteriol. 2001 May;183(10):3076-82. doi: 10.1128/JB.183.10.3076-3082.2001. J Bacteriol. 2001. PMID: 11325935 Free PMC article.
-
Role of the central region of NifL in conformational switches that regulate nitrogen fixation.Biochem Soc Trans. 2006 Feb;34(Pt 1):162-4. doi: 10.1042/BST0340162. Biochem Soc Trans. 2006. PMID: 16417511 Review.
Cited by
-
Genome-Wide Analyses of Proteome and Acetylome in Zymomonas mobilis Under N2-Fixing Condition.Front Microbiol. 2021 Oct 7;12:740555. doi: 10.3389/fmicb.2021.740555. eCollection 2021. Front Microbiol. 2021. PMID: 34803957 Free PMC article.
-
Complete genome sequence of Allochromatium vinosum DSM 180(T).Stand Genomic Sci. 2011 Dec 31;5(3):311-30. doi: 10.4056/sigs.2335270. Epub 2011 Dec 22. Stand Genomic Sci. 2011. PMID: 22675582 Free PMC article.
-
Nitrogen-Fixing Gamma Proteobacteria Azotobacter vinelandii-A Blueprint for Nitrogen-Fixing Plants?Microorganisms. 2024 Oct 18;12(10):2087. doi: 10.3390/microorganisms12102087. Microorganisms. 2024. PMID: 39458396 Free PMC article. Review.
-
Molecular Mechanism and Agricultural Application of the NifA-NifL System for Nitrogen Fixation.Int J Mol Sci. 2023 Jan 4;24(2):907. doi: 10.3390/ijms24020907. Int J Mol Sci. 2023. PMID: 36674420 Free PMC article. Review.
-
Diazotrophic Growth Allows Azotobacter vinelandii To Overcome the Deleterious Effects of a glnE Deletion.Appl Environ Microbiol. 2017 Jun 16;83(13):e00808-17. doi: 10.1128/AEM.00808-17. Print 2017 Jul 1. Appl Environ Microbiol. 2017. PMID: 28432097 Free PMC article.
References
-
- Schmitz, R. A., Klopprogge, K. & Grabbe, R. (2002) J. Mol. Microbiol. Biotechnol. 4, 235-242. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
