

RGS9-2 modulates D2 dopamine receptor-mediated Ca2+ channel inhibition in rat striatal cholinergic interneurons
- PMID: 15534226
- PMCID: PMC528982
- DOI: 10.1073/pnas.0407416101
RGS9-2 modulates D2 dopamine receptor-mediated Ca2+ channel inhibition in rat striatal cholinergic interneurons
Abstract
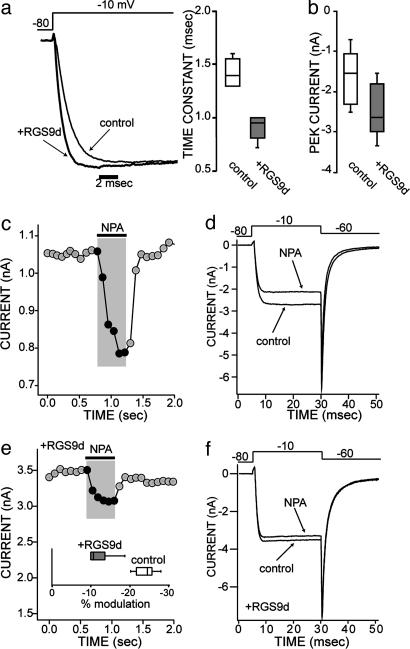
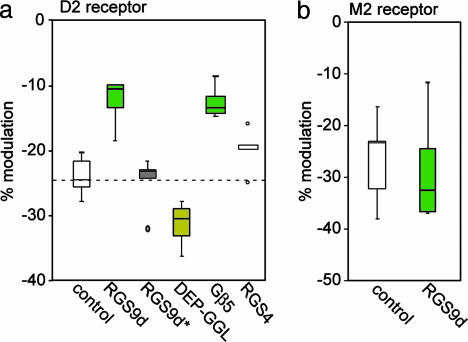
Regulator of G protein signaling (RGS) proteins negatively regulate receptor-mediated second messenger responses by enhancing the GTPase activity of Galpha subunits. We describe a receptor-specific role for an RGS protein at the level of an individual brain neuron. RGS9-2 and Gbeta(5) mRNA and protein complexes were detected in striatal cholinergic and gamma-aminobutyric acidergic neurons. Dialysis of cholinergic neurons with RGS9 constructs enhanced basal Ca(2+) channel currents and reduced D(2) dopamine receptor modulation of Cav2.2 channels. These constructs did not alter M(2) muscarinic receptor modulation of Cav2.2 currents in the same neuron. The noncatalytic DEP-GGL domain of RGS9 antagonized endogenous RGS9-2 activity, enhancing D(2) receptor modulation of Ca(2+) currents. In vitro, RGS9 constructs accelerated GTPase activity, in agreement with electrophysiological measurements, and did so more effectively at Go than Gi. These results implicate RGS9-2 as a specific regulator of dopamine receptor-mediated signaling in the striatum and identify a role for GAP activity modulation by the DEP-GGL domain.
Figures
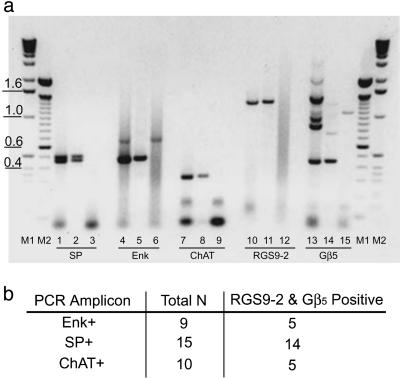
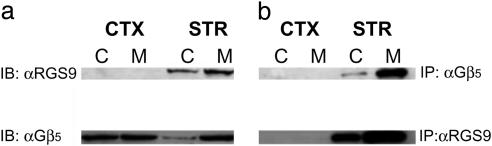
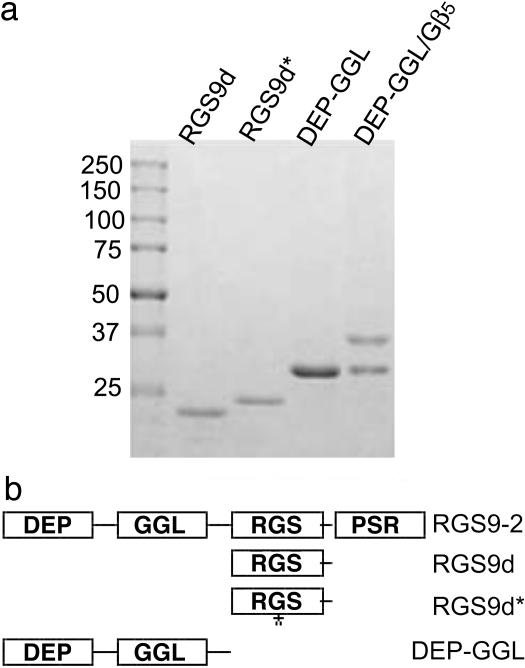
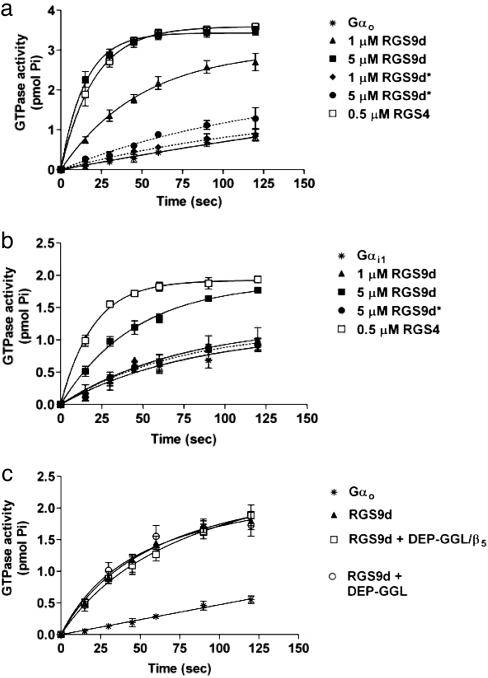
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 NS034696/NS/NINDS NIH HHS/United States
- EY10291/EY/NEI NIH HHS/United States
- R01 DA012958/DA/NIDA NIH HHS/United States
- 1F32NS10955-01/NS/NINDS NIH HHS/United States
- NS34696/NS/NINDS NIH HHS/United States
- EY06062/EY/NEI NIH HHS/United States
- R37 NS034696/NS/NINDS NIH HHS/United States
- R01 EY010291/EY/NEI NIH HHS/United States
- R01 EY006062/EY/NEI NIH HHS/United States
- T32 CA070085/CA/NCI NIH HHS/United States
- 5T32CA70085/CA/NCI NIH HHS/United States
- DA12958/DA/NIDA NIH HHS/United States
- F32 NS010955/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
