

Invertebrate carcasses as a resource for competing Aedes albopictus and Aedes aegypti (Diptera: Culicidae)
- PMID: 15535579
- PMCID: PMC2579927
- DOI: 10.1093/jmedent/37.3.364
Invertebrate carcasses as a resource for competing Aedes albopictus and Aedes aegypti (Diptera: Culicidae)
Abstract
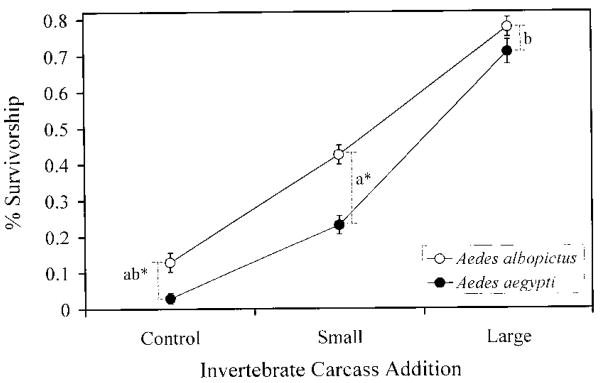
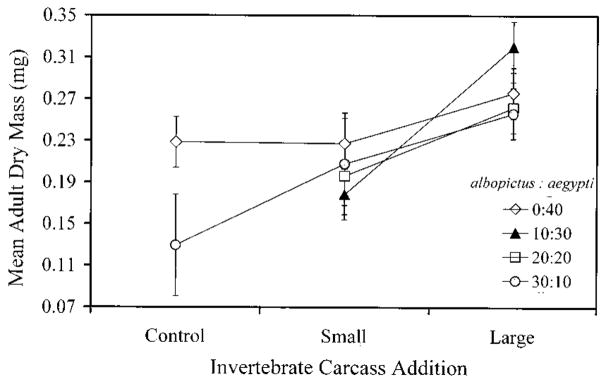
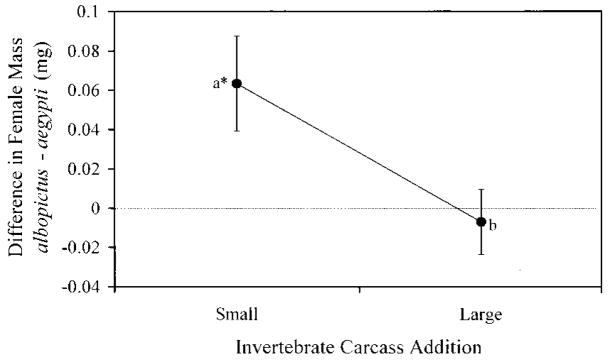
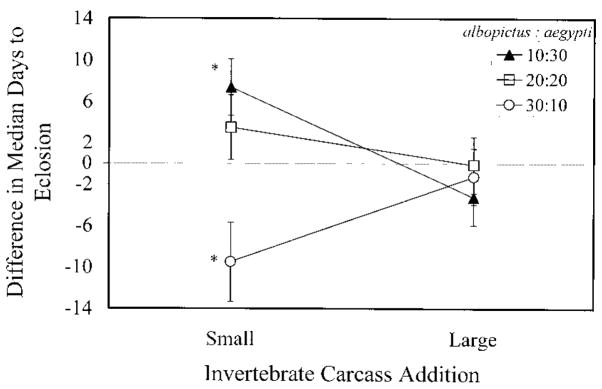
Terrestrial invertebrate carcasses are an important resource for insects developing in pitcher plants. However, little is known of the role of these carcasses in other containers, which also receive leaf fall and stemflow inputs. This experiment investigated effects of accumulated invertebrate carcasses as a resource for two competing mosquitoes, Aedes albopictus (Skuse) and Aedes aegypti (L.), whether either species differentially benefited from accumulated carcasses, and if such a benefit affected interspecific competition. First, we measured accumulation of invertebrate carcasses in standard containers at a field site. We then used a replacement series with five different species ratios at the same total density, and varied the input of invertebrate carcasses [dead Drosophila melanogaster (Meigen) ] in three levels: none, the average input from our field site, or the maximum input recorded at our field site. Survivorship, development time, and mass were measured for each mosquito species as correlates of population growth, and were used to calculate a population performance index, lambda'. There were strong positive effects of invertebrate carcass additions on all growth correlates and lambda'. Differences in performance between species were pronounced in small or no carcass additions and absent in large inputs of invertebrate carcasses, but there was little evidence that inputs of invertebrate carcasses altered the competitive advantage in this system. These results suggest that terrestrial invertebrate carcasses may be an important resource for many types of container communities, and large accumulations of dead invertebrates may reduce resource competition between these mosquitoes, thus favoring coexistence. We propose that the total amount of resource, including accumulated invertebrate carcasses, may explain observed patterns of replacement involving these mosquitoes.
Figures

References
-
- Barrera R. Competition and resistance to starvation in larvae of container-inhabiting Aedes mosquitoes. Ecol Entomol. 1996;21:117–127.
-
- Begon M, Harper JL, Townsend CR. Ecology. Blackwell; Oxford: 1990.
-
- Bradshaw WE. Interaction between the mosquito Wyeomia smithii, the midge Metriocnemus knabi, and their carnivorous host Sarracenia purpurea, In: Frank JH, Lounibos LP, editors. Phytotelmata: terrestrial plants as hosts for aquatic insect communities. Plexus; Medford, NJ: 1983. pp. 161–190.
-
- Bradshaw WE, Holzapfel CM. The evolution of insect life cycles. Springer; New York: 1986. Geography of density-dependent selection in pitcher-plant mosquitoes; pp. 48–65.
-
- Carpenter SR. Stemflow chemistry: effects on population dynamics of detritivorous mosquitoes in tree-hole ecosystems. Oecologia (Berl) 1982;53:1–6. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
