

Phosphorylation of 1-aminocyclopropane-1-carboxylic acid synthase by MPK6, a stress-responsive mitogen-activated protein kinase, induces ethylene biosynthesis in Arabidopsis
- PMID: 15539472
- PMCID: PMC535880
- DOI: 10.1105/tpc.104.026609
Phosphorylation of 1-aminocyclopropane-1-carboxylic acid synthase by MPK6, a stress-responsive mitogen-activated protein kinase, induces ethylene biosynthesis in Arabidopsis
Abstract
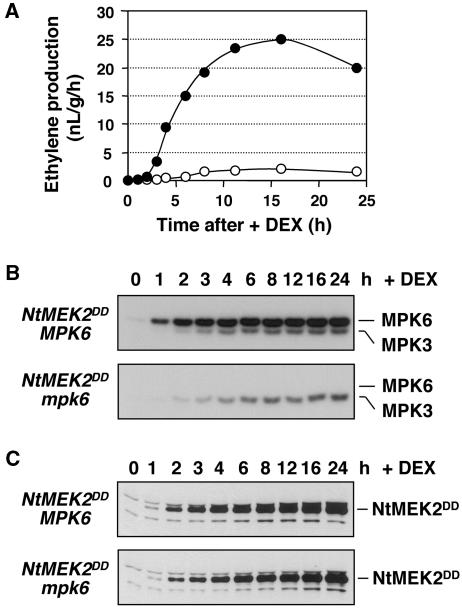
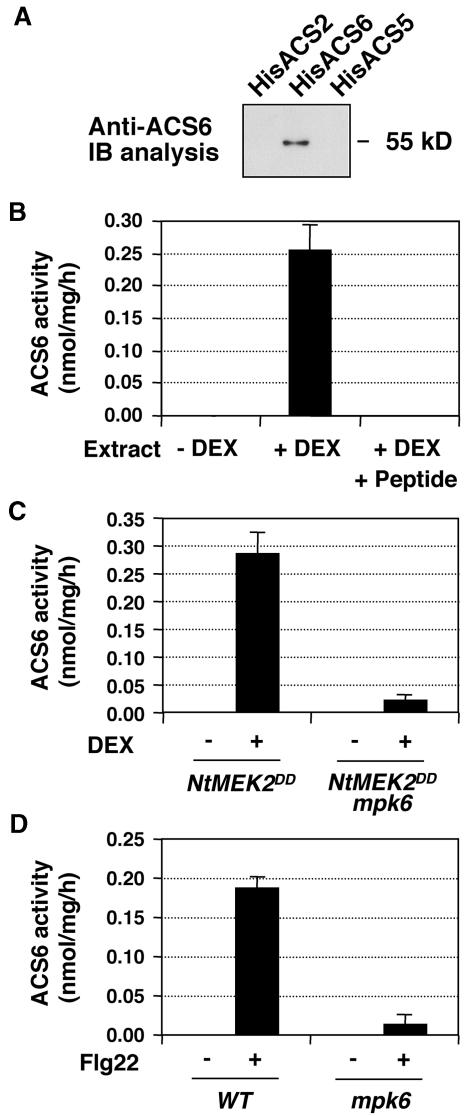
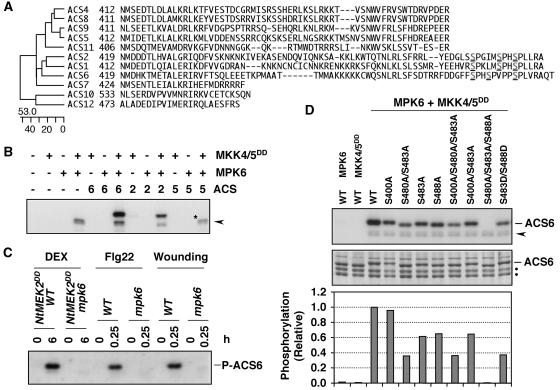
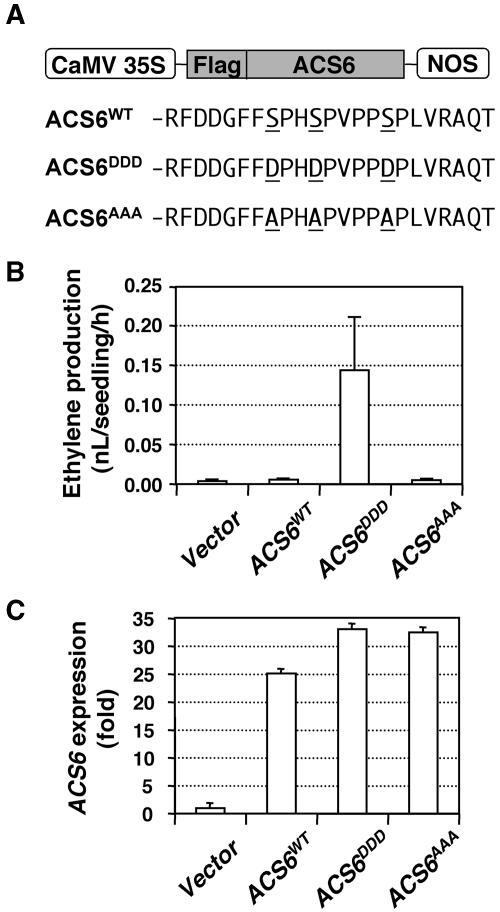
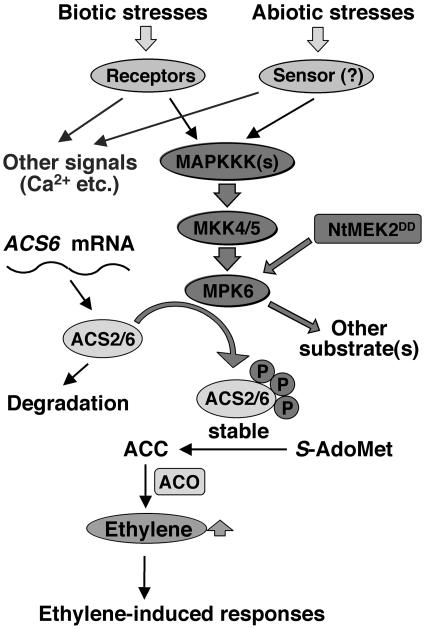
Mitogen-activated protein kinases (MAPKs) are implicated in regulating plant growth, development, and response to the environment. However, the underlying mechanisms are unknown because of the lack of information about their substrates. Using a conditional gain-of-function transgenic system, we demonstrated that the activation of SIPK, a tobacco (Nicotiana tabacum) stress-responsive MAPK, induces the biosynthesis of ethylene. Here, we report that MPK6, the Arabidopsis thaliana ortholog of tobacco SIPK, is required for ethylene induction in this transgenic system. Furthermore, we found that selected isoforms of 1-aminocyclopropane-1-carboxylic acid synthase (ACS), the rate-limiting enzyme of ethylene biosynthesis, are substrates of MPK6. Phosphorylation of ACS2 and ACS6 by MPK6 leads to the accumulation of ACS protein and, thus, elevated levels of cellular ACS activity and ethylene production. Expression of ACS6(DDD), a gain-of-function ACS6 mutant that mimics the phosphorylated form of ACS6, confers constitutive ethylene production and ethylene-induced phenotypes. Increasing numbers of stress stimuli have been shown to activate Arabidopsis MPK6 or its orthologs in other plant species. The identification of the first plant MAPK substrate in this report reveals one mechanism by which MPK6/SIPK regulates plant stress responses. Equally important, this study uncovers a signaling pathway that modulates the biosynthesis of ethylene, an important plant hormone, in plants under stress.
Figures



References
-
- Abeles, F.B., Morgan, P.W., and Saltveit, M.E.J. (1992). Ethylene in Plant Biology. (San Diego: Academic Press).
-
- Alonso, J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301, 653–657. - PubMed
-
- Aoyama, T., and Chua, N.-H. (1997). A glucocorticoid-mediated mediated transcriptional induction system in transgenic plants. Plant J. 11, 605–612. - PubMed
-
- Asai, T., Tena, G., Plotnikova, J., Willmann, M.R., Chiu, W.-L., Gomez-Gomez, L., Boller, T., Ausubel, F.M., and Sheen, J. (2002). MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 415, 977–983. - PubMed
-
- Bleecker, A.B., and Kende, H. (2000). Ethylene: A gaseous signal molecule in plants. Annu. Rev. Cell Dev. Biol. 16, 1–18. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
