

Bracoviruses contain a large multigene family coding for protein tyrosine phosphatases
- PMID: 15542661
- PMCID: PMC524979
- DOI: 10.1128/JVI.78.23.13090-13103.2004
Bracoviruses contain a large multigene family coding for protein tyrosine phosphatases
Abstract
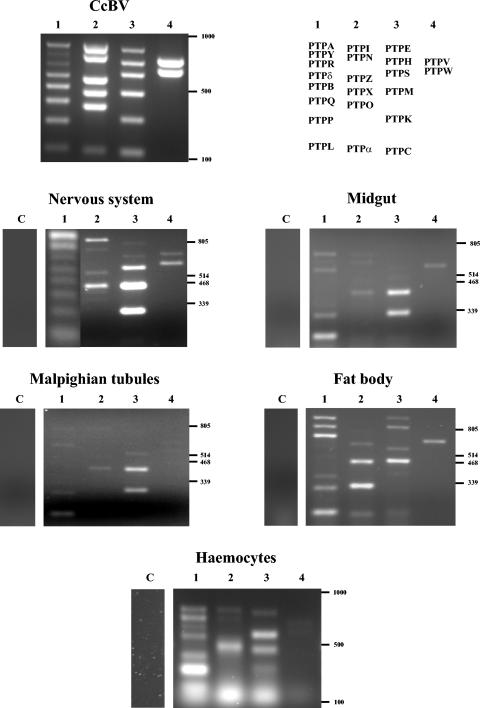
The relationship between parasitic wasps and bracoviruses constitutes one of the few known mutualisms between viruses and eukaryotes. The virions produced in the wasp ovaries are injected into host lepidopteran larvae, where virus genes are expressed, allowing successful development of the parasite by inducing host immune suppression and developmental arrest. Bracovirus-bearing wasps have a common phylogenetic origin, and contemporary bracoviruses are hypothesized to have been inherited by chromosomal transmission from a virus that originally integrated into the genome of the common ancestor wasp living 73.7 +/- 10 million years ago. However, so far no conserved genes have been described among different braconid wasp subfamilies. Here we show that a gene family is present in bracoviruses of different braconid wasp subfamilies (Cotesia congregata, Microgastrinae, and Toxoneuron nigriceps, Cardiochilinae) which likely corresponds to an ancient component of the bracovirus genome that might have been present in the ancestral virus. The genes encode proteins belonging to the protein tyrosine phosphatase family, known to play a key role in the control of signal transduction pathways. Bracovirus protein tyrosine phosphatase genes were shown to be expressed in different tissues of parasitized hosts, and two protein tyrosine phosphatases were produced with recombinant baculoviruses and tested for their biochemical activity. One protein tyrosine phosphatase is a functional phosphatase. These results strengthen the hypothesis that protein tyrosine phosphatases are involved in virally induced alterations of host physiology during parasitism.
Figures






Similar articles
-
Functional annotation of Cotesia congregata bracovirus: identification of viral genes expressed in parasitized host immune tissues.J Virol. 2014 Aug;88(16):8795-812. doi: 10.1128/JVI.00209-14. Epub 2014 May 28. J Virol. 2014. PMID: 24872581 Free PMC article.
-
The Complete Genome of Chelonus insularis Reveals Dynamic Arrangement of Genome Components in Parasitoid Wasps That Produce Bracoviruses.J Virol. 2022 Mar 9;96(5):e0157321. doi: 10.1128/JVI.01573-21. Epub 2022 Jan 5. J Virol. 2022. PMID: 34985997 Free PMC article.
-
Genome-Wide Patterns of Bracovirus Chromosomal Integration into Multiple Host Tissues during Parasitism.J Virol. 2021 Oct 27;95(22):e0068421. doi: 10.1128/JVI.00684-21. Epub 2021 Jul 28. J Virol. 2021. PMID: 34319152 Free PMC article.
-
Origin and evolution of polydnaviruses by symbiogenesis of insect DNA viruses in endoparasitic wasps.J Insect Physiol. 2003 May;49(5):419-32. doi: 10.1016/s0022-1910(03)00059-3. J Insect Physiol. 2003. PMID: 12770621 Review.
-
Polydnavirus hidden face: the genes producing virus particles of parasitic wasps.J Invertebr Pathol. 2009 Jul;101(3):194-203. doi: 10.1016/j.jip.2009.04.006. Epub 2009 May 19. J Invertebr Pathol. 2009. PMID: 19460382 Review.
Cited by
-
Characterizing virulence differences in a parasitoid wasp through comparative transcriptomic and proteomic.BMC Genomics. 2024 Oct 7;25(1):940. doi: 10.1186/s12864-024-10694-4. BMC Genomics. 2024. PMID: 39375606 Free PMC article.
-
Transient expression of protein tyrosine phosphatases encoded in Cotesia plutellae bracovirus inhibits insect cellular immune responses.Naturwissenschaften. 2008 Jan;95(1):25-32. doi: 10.1007/s00114-007-0290-7. Epub 2007 Jul 24. Naturwissenschaften. 2008. PMID: 17646950
-
Viral cystatin evolution and three-dimensional structure modelling: a case of directional selection acting on a viral protein involved in a host-parasitoid interaction.BMC Biol. 2008 Sep 10;6:38. doi: 10.1186/1741-7007-6-38. BMC Biol. 2008. PMID: 18783611 Free PMC article.
-
Functional annotation of Cotesia congregata bracovirus: identification of viral genes expressed in parasitized host immune tissues.J Virol. 2014 Aug;88(16):8795-812. doi: 10.1128/JVI.00209-14. Epub 2014 May 28. J Virol. 2014. PMID: 24872581 Free PMC article.
-
Convergent use of RhoGAP toxins by eukaryotic parasites and bacterial pathogens.PLoS Pathog. 2007 Dec 28;3(12):e203. doi: 10.1371/journal.ppat.0030203. PLoS Pathog. 2007. PMID: 18166080 Free PMC article.
References
-
- Albrecht, U., T. Wyler, R. Pfister-Wilhelm, A. Gruber, P. Stettler, P. Heiniger, E. Kurt, D. Schumperli, and B. Lanzrein. 1994. Polydnavirus of the parasitic wasp Chelonus inanitus (Braconidae): characterization, genome organization and time point of replication. J. Gen. Virol. 75:3353-3363. - PubMed
-
- Barford, D., A. J. Flint, and N. K. Tonks. 1994. Crystal structure of human protein tyrosine phosphatase 1B. Science 263:1397-1404. - PubMed
-
- Beckage, N. E., and I. De Buron. 1993. Lack of prothoracic gland degeneration in developmentally arrested host larvae of Manduca sexta parasitized by the braconid wasp Cotesia congregata. J. Invertebr. Pathol. 61:103-106.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
