

Dependence of ORC silencing function on NatA-mediated Nalpha acetylation in Saccharomyces cerevisiae
- PMID: 15542839
- PMCID: PMC529049
- DOI: 10.1128/MCB.24.23.10300-10312.2004
Dependence of ORC silencing function on NatA-mediated Nalpha acetylation in Saccharomyces cerevisiae
Abstract
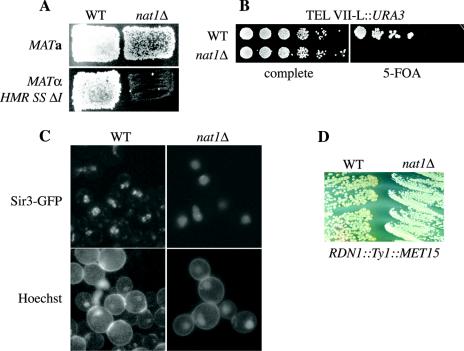
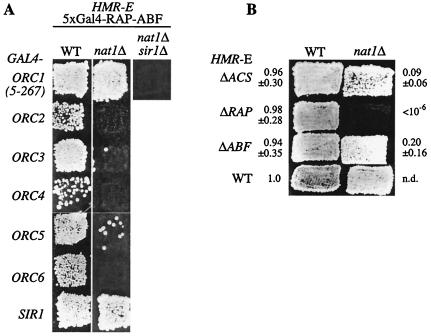
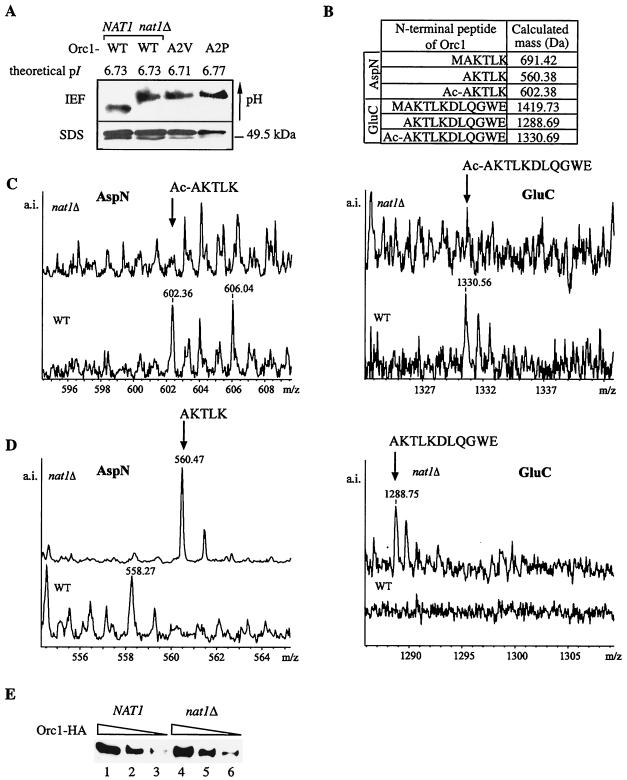
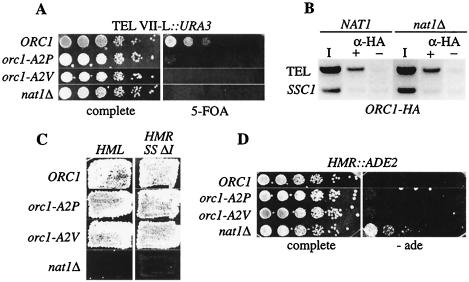
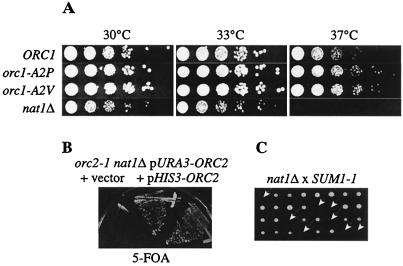
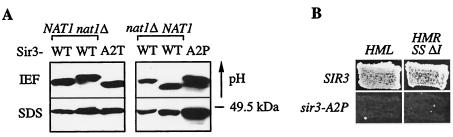
N(alpha) acetylation is one of the most abundant protein modifications in eukaryotes and is catalyzed by N-terminal acetyltransferases (NATs). NatA, the major NAT in Saccharomyces cerevisiae, consists of the subunits Nat1p, Ard1p, and Nat5p and is necessary for the assembly of repressive chromatin structures. Here, we found that Orc1p, the large subunit of the origin recognition complex (ORC), required NatA acetylation for its role in telomeric silencing. NatA functioned genetically through the ORC binding site of the HMR-E silencer. Furthermore, tethering Orc1p directly to the silencer circumvented the requirement for NatA in silencing. Orc1p was N(alpha) acetylated in vivo by NatA. Mutations that abrogated its ability to be acetylated caused strong telomeric derepression. Thus, N(alpha) acetylation of Orc1p represents a protein modification that modulates chromatin function in S. cerevisiae. Genetic evidence further supported a functional link between NatA and ORC: (i) nat1Delta was synthetically lethal with orc2-1 and (ii) the synthetic lethality between nat1Delta and SUM1-1 required the Orc1 N terminus. We also found Sir3p to be acetylated by NatA. In summary, we propose a model by which N(alpha) acetylation is required for the binding of silencing factors to the N terminus of Orc1p and Sir3p to recruit heterochromatic factors and establish repression.
Figures
Similar articles
-
Functions of protosilencers in the formation and maintenance of heterochromatin in Saccharomyces cerevisiae.PLoS One. 2012;7(5):e37092. doi: 10.1371/journal.pone.0037092. Epub 2012 May 17. PLoS One. 2012. PMID: 22615905 Free PMC article.
-
Importance of the Sir3 N terminus and its acetylation for yeast transcriptional silencing.Genetics. 2004 Sep;168(1):547-51. doi: 10.1534/genetics.104.028803. Genetics. 2004. PMID: 15454564 Free PMC article.
-
A region of the nucleosome required for multiple types of transcriptional silencing in Saccharomyces cerevisiae.Genetics. 2011 Jul;188(3):535-48. doi: 10.1534/genetics.111.129197. Epub 2011 May 5. Genetics. 2011. PMID: 21546544 Free PMC article.
-
N-terminal acetyltransferases and sequence requirements for N-terminal acetylation of eukaryotic proteins.J Mol Biol. 2003 Jan 24;325(4):595-622. doi: 10.1016/s0022-2836(02)01269-x. J Mol Biol. 2003. PMID: 12507466 Review.
-
Silent information regulator 3: the Goldilocks of the silencing complex.Genes Dev. 2010 Jan 15;24(2):115-22. doi: 10.1101/gad.1865510. Genes Dev. 2010. PMID: 20080949 Free PMC article. Review.
Cited by
-
Structure and function of the Saccharomyces cerevisiae Sir3 BAH domain.Mol Cell Biol. 2006 Apr;26(8):3256-65. doi: 10.1128/MCB.26.8.3256-3265.2006. Mol Cell Biol. 2006. PMID: 16581798 Free PMC article.
-
Trypanosoma brucei Orc1 is essential for nuclear DNA replication and affects both VSG silencing and VSG switching.Mol Microbiol. 2013 Jan;87(1):196-210. doi: 10.1111/mmi.12093. Epub 2012 Dec 10. Mol Microbiol. 2013. PMID: 23216794 Free PMC article.
-
Physiological importance and identification of novel targets for the N-terminal acetyltransferase NatB.Eukaryot Cell. 2006 Feb;5(2):368-78. doi: 10.1128/EC.5.2.368-378.2006. Eukaryot Cell. 2006. PMID: 16467477 Free PMC article.
-
Characterization of hARD2, a processed hARD1 gene duplicate, encoding a human protein N-alpha-acetyltransferase.BMC Biochem. 2006 Apr 25;7:13. doi: 10.1186/1471-2091-7-13. BMC Biochem. 2006. PMID: 16638120 Free PMC article.
-
Clustering heterochromatin: Sir3 promotes telomere clustering independently of silencing in yeast.J Cell Biol. 2011 Feb 7;192(3):417-31. doi: 10.1083/jcb.201008007. J Cell Biol. 2011. PMID: 21300849 Free PMC article.
References
-
- Aparicio, O. M., B. L. Billington, and D. E. Gottschling. 1991. Modifiers of position effect are shared between telomeric and silent mating-type loci in Saccharomyces cerevisiae. Cell 66:1279-1287. - PubMed
-
- Arnold, R. J., B. Polevoda, J. P. Reilly, and F. Sherman. 1999. The action of N-terminal acetyltransferases on yeast ribosomal proteins. J. Biol. Chem. 274:37035-37040. - PubMed
-
- Bell, S. P., R. Kobayashi, and B. Stillman. 1993. Yeast origin recognition complex functions in transcription silencing and DNA replication. Science 262:1844-1849. - PubMed
-
- Bell, S. P., J. Mitchell, J. Leber, R. Kobayashi, and B. Stillman. 1995. The multidomain structure of Orc1p reveals similarity to regulators of DNA replication and transcriptional silencing. Cell 83:563-568. - PubMed
-
- Brand, A. H., G. Micklem, and K. Nasmyth. 1987. A yeast silencer contains sequences that can promote autonomous plasmid replication and transcriptional activation. Cell 51:709-719. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases