

Genetic interaction between Rb and K-ras in the control of differentiation and tumor suppression
- PMID: 15542848
- PMCID: PMC529028
- DOI: 10.1128/MCB.24.23.10406-10415.2004
Genetic interaction between Rb and K-ras in the control of differentiation and tumor suppression
Abstract
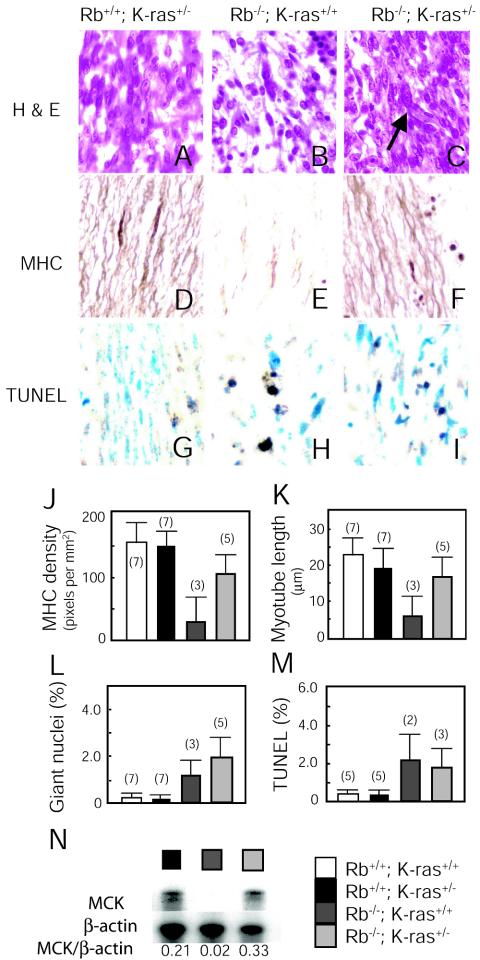
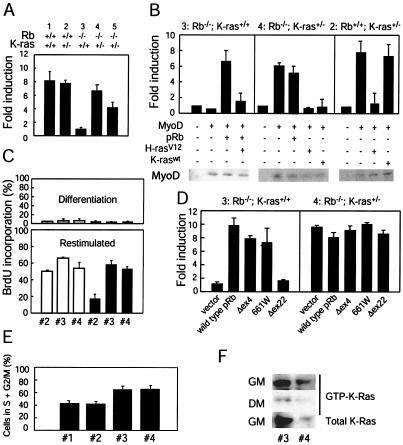
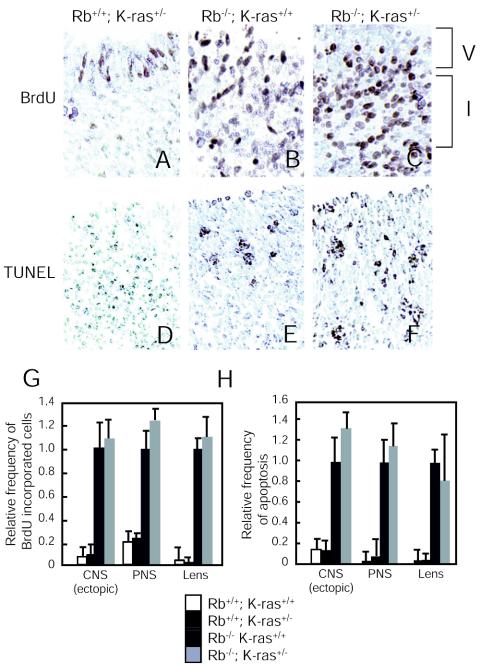
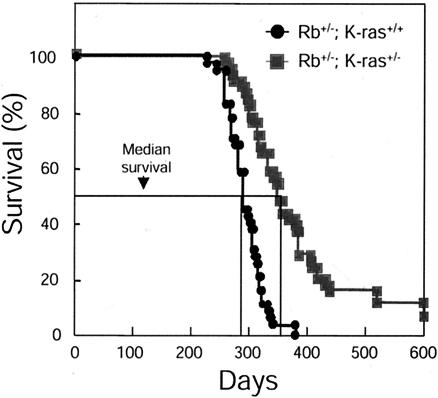
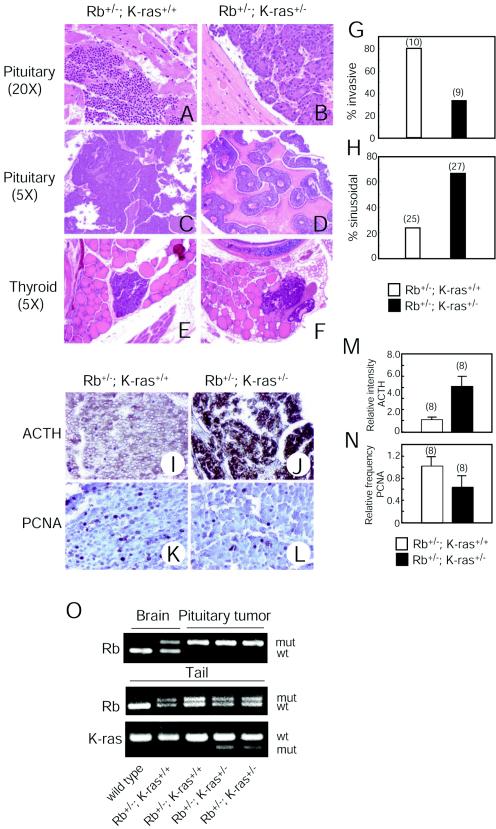
Although the retinoblastoma protein (pRb) has been implicated in the processes of cellular differentiation, there is no compelling genetic or in vivo evidence that such activities contribute to pRb-mediated tumor suppression. Motivated by cell culture studies suggesting that Ras is a downstream effector of pRb in the control of differentiation, we have examined the tumor and developmental phenotypes of Rb and K-ras double-knockout mice. We find that heterozygosity for K-ras (i) rescued a unique subset of developmental defects that characterize Rb-deficient embryos by affecting differentiation but not proliferation and (ii) significantly enhanced the degree of differentiation of pituitary adenocarcinomas arising in Rb heterozygotes, leading to their prolonged survival. These observations suggest that Rb and K-ras function together in vivo, in the contexts of both embryonic and tumor development, and that the ability to affect differentiation is a major facet of the tumor suppressor function of pRb.
Figures
Similar articles
-
Rb and N-ras function together to control differentiation in the mouse.Mol Cell Biol. 2003 Aug;23(15):5256-68. doi: 10.1128/MCB.23.15.5256-5268.2003. Mol Cell Biol. 2003. PMID: 12861012 Free PMC article.
-
Genetic interaction between Rb and N-ras: differentiation control and metastasis.Cancer Res. 2006 Oct 1;66(19):9345-8. doi: 10.1158/0008-5472.CAN-06-1250. Cancer Res. 2006. PMID: 17018584 Review.
-
Activation of retinoblastoma protein in mammary gland leads to ductal growth suppression, precocious differentiation, and adenocarcinoma.J Cell Biol. 2002 Jan 7;156(1):185-98. doi: 10.1083/jcb.200106084. Epub 2002 Jan 3. J Cell Biol. 2002. PMID: 11777937 Free PMC article.
-
Id2 is a retinoblastoma protein target and mediates signalling by Myc oncoproteins.Nature. 2000 Oct 5;407(6804):592-8. doi: 10.1038/35036504. Nature. 2000. PMID: 11034201
-
[The retinoblastoma gene: from its basic understanding as a signal mediator for growth and differentiation to its use in the treatment of cancer].Gan To Kagaku Ryoho. 1997 Sep;24(11):1368-80. Gan To Kagaku Ryoho. 1997. PMID: 9309128 Review. Japanese.
Cited by
-
Wild-type NRas and KRas perform distinct functions during transformation.Mol Cell Biol. 2007 Oct;27(19):6742-55. doi: 10.1128/MCB.00234-07. Epub 2007 Jul 16. Mol Cell Biol. 2007. PMID: 17636015 Free PMC article.
-
Twists in views on RB functions in cellular signaling, metabolism and stem cells.Cancer Sci. 2012 Jul;103(7):1182-8. doi: 10.1111/j.1349-7006.2012.02284.x. Epub 2012 Apr 27. Cancer Sci. 2012. PMID: 22448711 Free PMC article. Review.
-
Antitumor mechanisms when pRb and p53 are genetically inactivated.Oncogene. 2015 Aug 27;34(35):4547-57. doi: 10.1038/onc.2014.399. Epub 2014 Dec 8. Oncogene. 2015. PMID: 25486431 Free PMC article. Review.
-
The role of Cdk5 in neuroendocrine thyroid cancer.Cancer Cell. 2013 Oct 14;24(4):499-511. doi: 10.1016/j.ccr.2013.08.027. Cancer Cell. 2013. PMID: 24135281 Free PMC article.
-
p66(Shc) restrains Ras hyperactivation and suppresses metastatic behavior.Oncogene. 2010 Oct 14;29(41):5559-67. doi: 10.1038/onc.2010.326. Epub 2010 Aug 2. Oncogene. 2010. PMID: 20676142 Free PMC article.
References
-
- Capen, C. C., E. Karbe, U. Deschl, C. George, P.-G. Germann, C. Gopinath, J. F. Hardisty, J. Kanno, W. Kaufmann, G. Krinke, K. Kuttler, B. Kulwich, C. Landes, B. Lenz, L. Longeart, I. Paulson, E. Sander, and K. Tuch. 2001. Endocrine system, p. 269-322. In U. Mohr (ed.), International classification of rodent tumors: the mouse. Springer-Verlag, Berlin, Germany.
-
- Charles, A., X. Tang, E. Crouch, J. S. Brody, and Z. X. Xiao. 2001. Retinoblastoma protein complexes with C/EBP proteins and activates C/EBP-mediated transcription. J. Cell. Biochem. 83:414-425. - PubMed
-
- Chen, P.-L., D. J. Riley, Y. Chen, and W.-H. Lee. 1996. Retinoblastoma protein positively regulates terminal adipocyte differentiation through direct interaction with C/EBPs. Genes Dev. 10:2794-2804. - PubMed
-
- Clarke, A. R., E. R. Maandag, M. van Roon, N. M. T. van der Lugt, M. van der Valk, M. L. Hooper, A. Berns, and H. te Riele. 1992. Requirement for a functional Rb-1 gene in murine development. Nature 359:328-330. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous