

Sox7 plays crucial roles in parietal endoderm differentiation in F9 embryonal carcinoma cells through regulating Gata-4 and Gata-6 expression
- PMID: 15542856
- PMCID: PMC529033
- DOI: 10.1128/MCB.24.23.10492-10503.2004
Sox7 plays crucial roles in parietal endoderm differentiation in F9 embryonal carcinoma cells through regulating Gata-4 and Gata-6 expression
Abstract
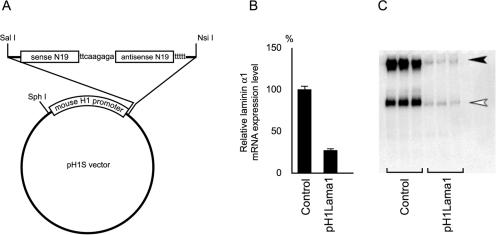
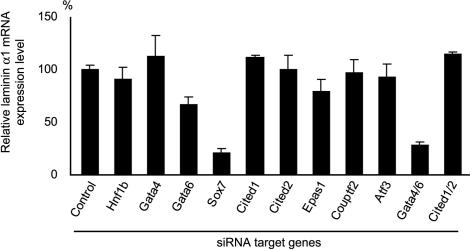
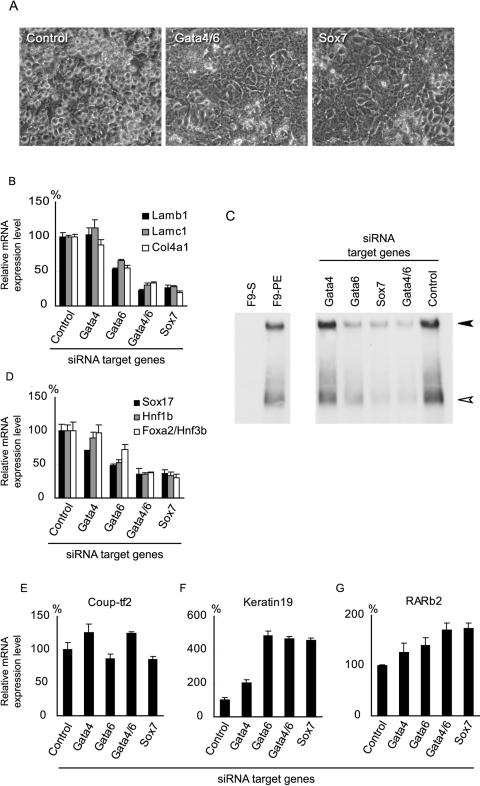
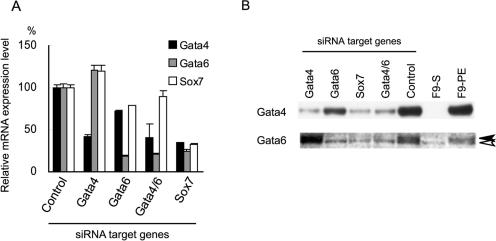
During early rodent development, the parietal endoderm appears from an inner cell mass and produces large amounts of basement membrane components, such as laminin-1 and collagen IV. To elucidate the regulatory network for gene expression during these procedures, we constructed a series of short interfering RNA expression vectors targeted to various transcription factors, transfected them into F9 embryonal carcinoma cells, and evaluated the effects of the gene silencing on the induction of parietal endoderm differentiation and basement membrane component production by treating F9 cells with all trans-retinoic acid and dibutyryl cyclic AMP. Among the transcription factors tested, silencing of Sox7 or combined silencing of Gata-4 and Gata-6 resulted in suppression of cell shape changes and laminin-1 production, which are the hallmarks of parietal endoderm differentiation. In cells silenced for Sox7, induction of Gata-4 and Gata-6 by retinoic acid and cyclic AMP treatment was inhibited, while induction of Sox7 was not affected in cells silenced for Gata-4 and Gata-6, indicating that Sox7 is an upstream regulatory factor for these Gata factors. Nevertheless, silencing of Sox7 did not totally cancel the action of retinoic acid, since upregulation of coup-tf2, keratin 19, and retinoic acid receptor beta2 was not abolished in Sox7-silenced F9 cells. Although overexpression of Sox7 alone was insufficient to induce parietal endoderm differentiation, overexpression of Gata-4 or Gata-6 in Sox7-silenced F9 cells restored the differentiation into parietal endoderm. Sox7 is therefore required for the induction of Gata-4 and Gata-6, and the interplay among these transcription factors plays a crucial role in parietal endoderm differentiation.
Figures
Similar articles
-
SOX7 and GATA-4 are competitive activators of Fgf-3 transcription.J Biol Chem. 2004 Jul 2;279(27):28564-73. doi: 10.1074/jbc.M313814200. Epub 2004 Apr 13. J Biol Chem. 2004. PMID: 15082719
-
Differentiation of embryonic stem cells is induced by GATA factors.Genes Dev. 2002 Apr 1;16(7):784-9. doi: 10.1101/gad.968802. Genes Dev. 2002. PMID: 11937486 Free PMC article.
-
SOX7 and SOX17 regulate the parietal endoderm-specific enhancer activity of mouse laminin alpha1 gene.J Biol Chem. 2004 Sep 3;279(36):38055-61. doi: 10.1074/jbc.M403724200. Epub 2004 Jun 25. J Biol Chem. 2004. PMID: 15220343
-
Role of GATA factors in development, differentiation, and homeostasis of the small intestinal epithelium.Am J Physiol Gastrointest Liver Physiol. 2014 Mar;306(6):G474-90. doi: 10.1152/ajpgi.00119.2013. Epub 2014 Jan 16. Am J Physiol Gastrointest Liver Physiol. 2014. PMID: 24436352 Free PMC article. Review.
-
Sox7 in vascular development: review, insights and potential mechanisms.Int J Dev Biol. 2014;58(1):1-8. doi: 10.1387/ijdb.130323mw. Int J Dev Biol. 2014. PMID: 24860989 Review.
Cited by
-
De novo copy number variants are associated with congenital diaphragmatic hernia.J Med Genet. 2012 Oct;49(10):650-9. doi: 10.1136/jmedgenet-2012-101135. J Med Genet. 2012. PMID: 23054247 Free PMC article.
-
Serpent/dGATAb regulates Laminin B1 and Laminin B2 expression during Drosophila embryogenesis.Sci Rep. 2019 Nov 4;9(1):15910. doi: 10.1038/s41598-019-52210-9. Sci Rep. 2019. PMID: 31685844 Free PMC article.
-
Dissecting the retinoid-induced differentiation of F9 embryonal stem cells by integrative genomics.Mol Syst Biol. 2011 Oct 11;7:538. doi: 10.1038/msb.2011.73. Mol Syst Biol. 2011. PMID: 21988834 Free PMC article.
-
SOX7: from a developmental regulator to an emerging tumor suppressor.Histol Histopathol. 2014 Apr;29(4):439-45. doi: 10.14670/HH-29.10.439. Epub 2013 Nov 29. Histol Histopathol. 2014. PMID: 24288056 Free PMC article. Review.
-
The primitive endoderm lineage of the mouse blastocyst: sequential transcription factor activation and regulation of differentiation by Sox17.Dev Biol. 2011 Feb 15;350(2):393-404. doi: 10.1016/j.ydbio.2010.12.007. Epub 2010 Dec 10. Dev Biol. 2011. PMID: 21146513 Free PMC article.
References
-
- Alexander, J., and D. Y. Stainier. 1999. A molecular pathway leading to endoderm formation in zebrafish. Curr. Biol. 9:1147-1157. - PubMed
-
- Baldwin, C. T., J. E. Silbert, D. E. Humphries, J. N. Cogburn, and B. D. Smith. 1989. Increased proteoglycan synthesis following the differentiation of F9 embryonal carcinoma cells: formation of a differentiation-specific proteoheparan sulfate. Matrix 9:389-396. - PubMed
-
- Bamforth, S. D., J. Braganca, J. J. Eloranta, J. N. Murdoch, F. I. Marques, K. R. Kranc, H. Farza, D. J. Henderson, H. C. Hurst, and S. Bhattacharya. 2001. Cardiac malformations, adrenal agenesis, neural crest defects and exencephaly in mice lacking Cited2, a new Tfap2 co-activator. Nat. Genet. 29:469-474. - PubMed
-
- Barbacci, E., M. Reber, M. O. Ott, C. Breillat, F. Huetz, and S. Cereghini. 1999. Variant hepatocyte nuclear factor 1 is required for visceral endoderm specification. Development 126:4795-4805. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous