

Co-infection of malaria and gamma-herpesvirus: exacerbated lung inflammation or cross-protection depends on the stage of viral infection
- PMID: 15544614
- PMCID: PMC1809251
- DOI: 10.1111/j.1365-2249.2004.02652.x
Co-infection of malaria and gamma-herpesvirus: exacerbated lung inflammation or cross-protection depends on the stage of viral infection
Abstract
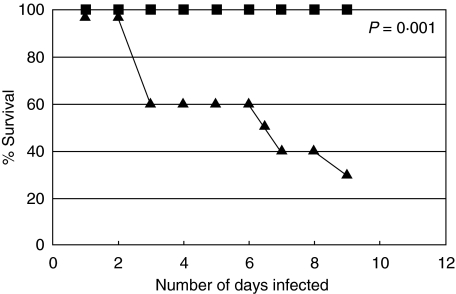
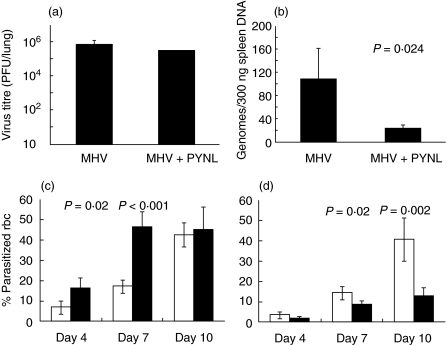
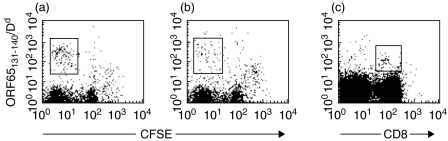
In order to study the interaction between a gamma-herpesvirus and malaria we established a co-infection model that involves infection of mice with murine gamma-herpesvirus (MHV-68) and Plasmodium yoelii non-lethal strain (PYNL). To investigate the interaction between acute malaria and the lytic stage of MHV-68, the timing of infections was chosen such that the peak virus and parasite burdens would be present at the same time. Under this condition, we observed significant mortality in co-infected mice and aggressive lung inflammation with a marked influx of neutrophils and megakaryocytes. If mice were latently infected with MHV-68 and then co-infected with malaria we noticed significantly less viral load and parasitaemia. Using MHC/peptide tetramer staining we found that acute malaria reduces the anti-MHV-68 CD8+ T cell response in the animals that develop severe disease. Our study provides important fundamental information, which will be of use when devising strategies to combat infections with more than one agent, a situation that often occurs naturally.
Figures


References
-
- Stiller CA, Parkin DM. Geographic and ethnic variations in the incidence of childhood cancer. Br Med Bull. 1996;52:682–703. - PubMed
-
- de The G. The etiology of Burkitt's lymphoma and the history of the shaken dogmas. Blood Cells. 1993;19:667–73. - PubMed
-
- de The G. Co-carcinogenic events in herpesvirus oncogenesis: a review. IARC Sci Publ. 1978:933–45. - PubMed
-
- Virgin HW, Speck SH. Unraveling immunity to gamma-herpesviruses: a new model for understanding the role of immunity in chronic virus infection. Curr Opin Immunol. 1999;11:371–9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
