

The RNA-binding protein SUP-12 controls muscle-specific splicing of the ADF/cofilin pre-mRNA in C. elegans
- PMID: 15545320
- PMCID: PMC1781344
- DOI: 10.1083/jcb.200407085
The RNA-binding protein SUP-12 controls muscle-specific splicing of the ADF/cofilin pre-mRNA in C. elegans
Abstract
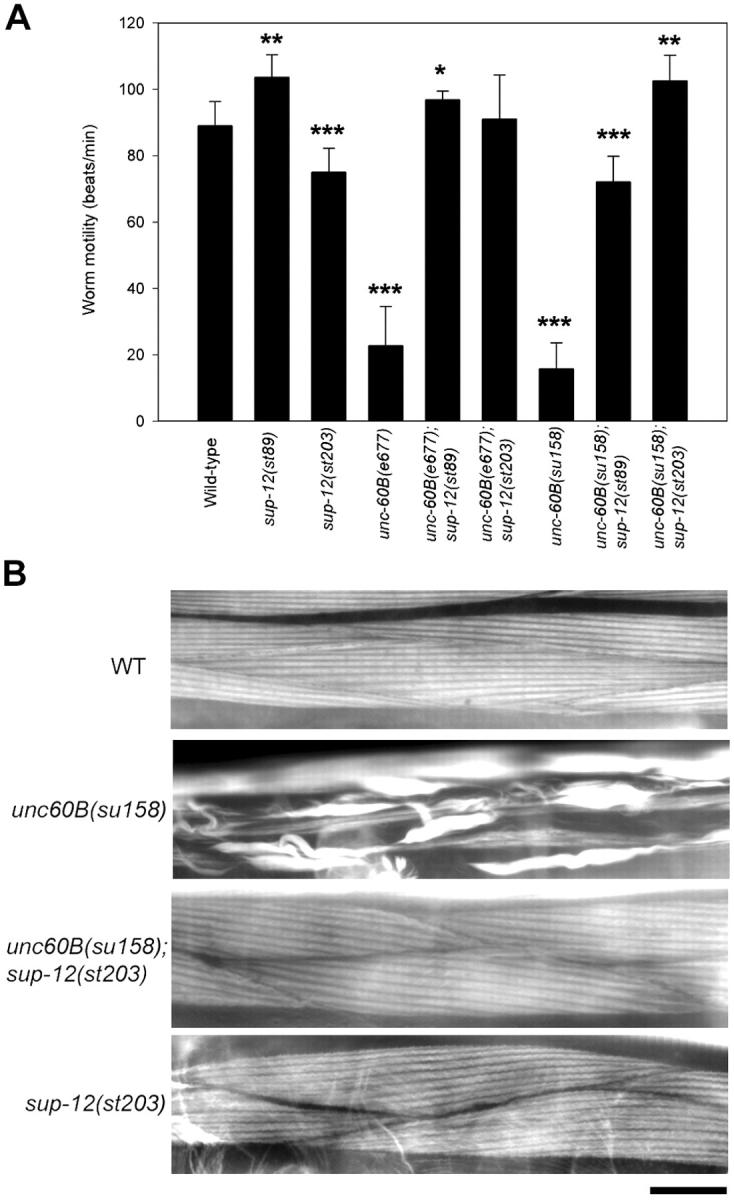
Tissue-specific alternative pre-mRNA splicing is essential for increasing diversity of functionally different gene products. In Caenorhabditis elegans, UNC-60A and UNC-60B, nonmuscle and muscle isoforms of actin depolymerizing factor (ADF)/cofilin, are expressed by alternative splicing of unc-60 and regulate distinct actin-dependent developmental processes. We report that SUP-12, a member of a new family of RNA recognition motif (RRM) proteins, including SEB-4, regulates muscle-specific splicing of unc-60. In sup-12 mutants, expression of UNC-60B is decreased, whereas UNC-60A is up-regulated in muscle. sup-12 mutations strongly suppress muscle defects in unc-60B mutants by allowing expression of UNC-60A in muscle that can substitute for UNC-60B, thus unmasking their functional redundancy. SUP-12 is expressed in muscle and localized to the nuclei in a speckled pattern. The RRM domain of SUP-12 binds to several sites of the unc-60 pre-mRNA including the UG repeats near the 3'-splice site in the first intron. Our results suggest that SUP-12 is a novel tissue-specific splicing factor and regulates functional redundancy among ADF/cofilin isoforms.
Figures





References
-
- Bamburg, J.R. 1999. Proteins of the ADF/cofilin family: essential regulators of actin dynamics. Annu. Rev. Cell Dev. Biol. 15:185–230. - PubMed
-
- Bamburg, J.R., A. McGough, and S. Ono. 1999. Putting a new twist on actin: ADF/cofilins modulate actin dynamics. Trends Cell Biol. 9:364–370. - PubMed
-
- Bentley, D. 2002. The mRNA assembly line: transcription and processing machines in the same factory. Curr. Opin. Cell Biol. 14:336–342. - PubMed
-
- Black, D.L. 2003. Mechanisms of alternative pre-messenger RNA splicing. Annu. Rev. Biochem. 72:291–336. - PubMed
-
- Boy, S., J. Souopgui, M.A. Amato, M. Wegnez, T. Pieler, and M. Perron. 2004. XSEB4R, a novel RNA-binding protein involved in retinal cell differentiation downstream of bHLH proneural genes. Development. 131:851–862. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
