

Determinants of human immunodeficiency virus type 1 escape from the primary CD8+ cytotoxic T lymphocyte response
- PMID: 15545352
- PMCID: PMC2211924
- DOI: 10.1084/jem.20040511
Determinants of human immunodeficiency virus type 1 escape from the primary CD8+ cytotoxic T lymphocyte response
Abstract
CD8+ cytotoxic T lymphocytes (CTLs) play an important role in containment of virus replication in primary human immunodeficiency virus (HIV) infection. HIV's ability to mutate to escape from CTL pressure is increasingly recognized; but comprehensive studies of escape from the CD8 T cell response in primary HIV infection are currently lacking. Here, we have fully characterized the primary CTL response to autologous virus Env, Gag, and Tat proteins in three patients, and investigated the extent, kinetics, and mechanisms of viral escape from epitope-specific components of the response. In all three individuals, we observed variation beginning within weeks of infection at epitope-containing sites in the viral quasispecies, which conferred escape by mechanisms including altered peptide presentation/recognition and altered antigen processing. The number of epitope-containing regions exhibiting evidence of early CTL escape ranged from 1 out of 21 in a subject who controlled viral replication effectively to 5 out of 7 in a subject who did not. Evaluation of the extent and kinetics of HIV-1 escape from >40 different epitope-specific CD8 T cell responses enabled analysis of factors determining escape and suggested that escape is restricted by costs to intrinsic viral fitness and by broad, codominant distribution of CTL-mediated pressure on viral replication.
Figures


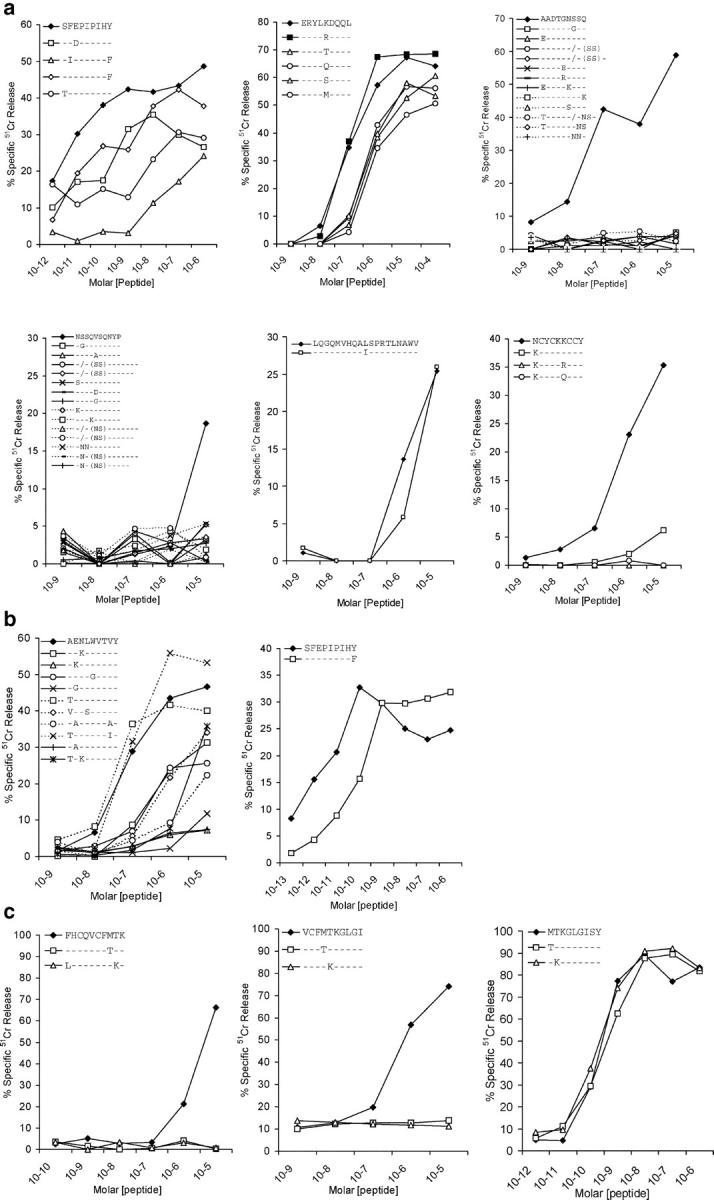
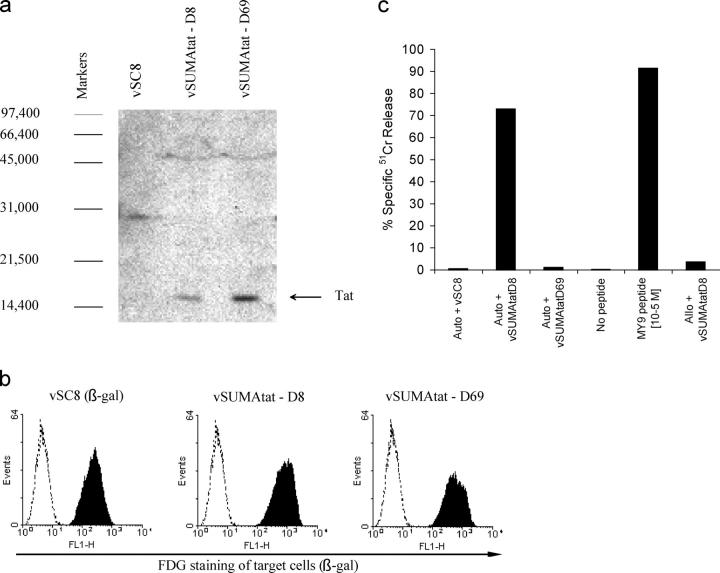
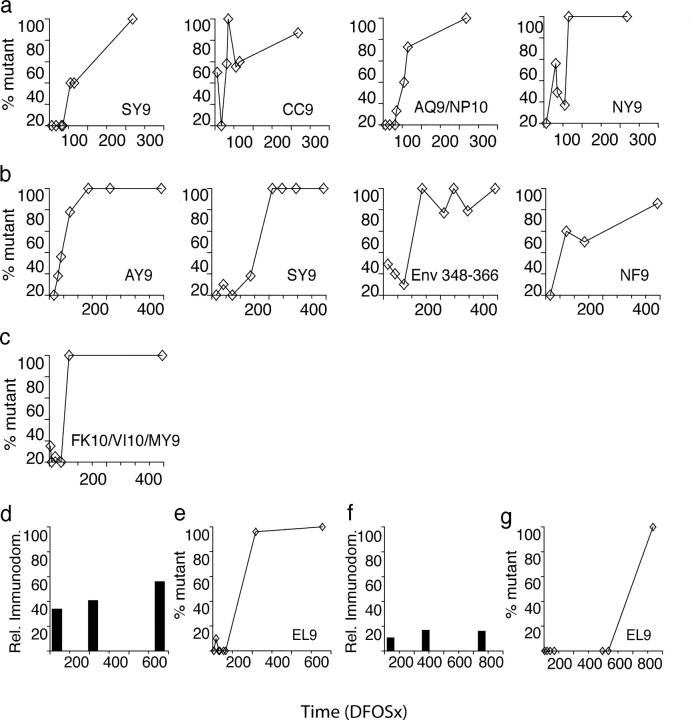
References
-
- McMichael, A.J., and S.L. Rowland-Jones. 2001. Cellular immune responses to HIV. Nature. 410:980–987. - PubMed
-
- McMichael, A., and T. Hanke. 2002. The quest for an AIDS vaccine: is the CD8+ T-cell approach feasible? Nat. Rev. Immunol. 2:283–291. - PubMed
-
- Barouch, D.H., and N.L. Letvin. 2002. Viral evolution and challenges in the development of HIV vaccines. Vaccine. 20:A66–A68. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
