

Sarco(endo)plasmic reticulum Ca2+ ATPase (SERCA) gene silencing and remodeling of the Ca2+ signaling mechanism in cardiac myocytes
- PMID: 15546997
- PMCID: PMC528906
- DOI: 10.1073/pnas.0407537101
Sarco(endo)plasmic reticulum Ca2+ ATPase (SERCA) gene silencing and remodeling of the Ca2+ signaling mechanism in cardiac myocytes
Abstract
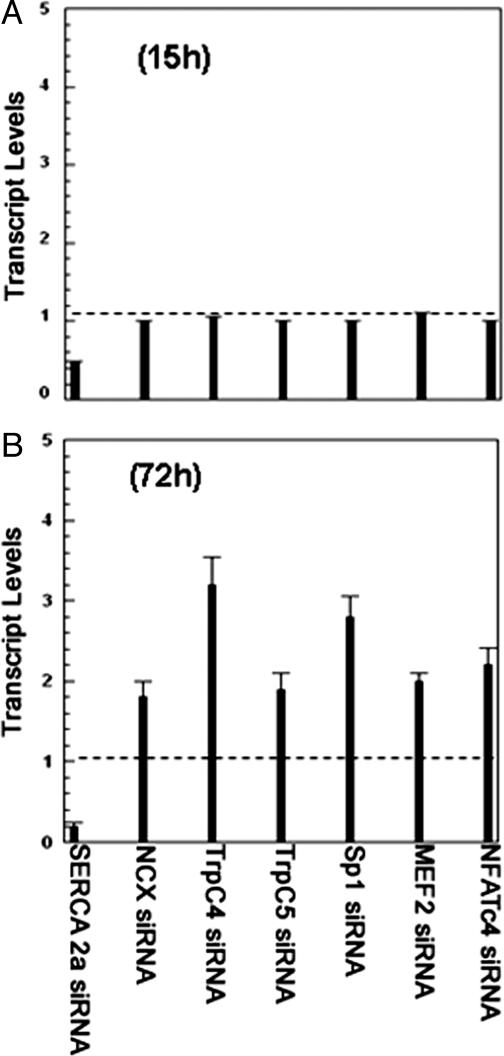
Transient elevations of cytosolic Ca2+ are a common mechanism of cellular signaling. In striated muscle, the sarco(endo)plasmic reticulum Ca2+ ATPase (SERCA) plays an important role in terminating Ca2+ transients by returning cytosolic Ca2+ to intracellular stores. Stored Ca2+ can then be released again for subsequent signaling. We down-regulated SERCA2 gene expression in cultured cardiac myocytes by means of endogenous transcription of small interfering RNA encoded by an exogenous cDNA template. The cDNA template was delivered by adenovirus vector. Reduction of SERCA expression in all myocytes in culture was documented by immunochemistry, real-time RT-PCR, and determination of ATP-dependent Ca2+ transport. The reduction of SERCA2 expression was associated with the up-regulation of transient receptor potential (TRP) channel proteins (TRPC4 and TRPC5) and Na+/Ca2+ exchanger, indicating that intracellular store deficiency was compensated for by Ca2+ fluxes through the plasma membrane. In fact, SERCA silencing was followed by increased transcription of Na+/Ca2+ exchanger, TRPC4, TRPC5, and related transcriptional factors, such as stimulating protein 1, myocyte enhancer factor 2, and nuclear factor of activated cells 4, through activation of calcineurin. This finding demonstrates that the observed compensation occurs through transcriptional crosstalk and the remodeling of Ca2+ signaling pathways. The wide significance of this regulatory mechanism is related to its general involvement in Ca2+ signaling dynamics and in cardiac development and hypertrophy.
Figures
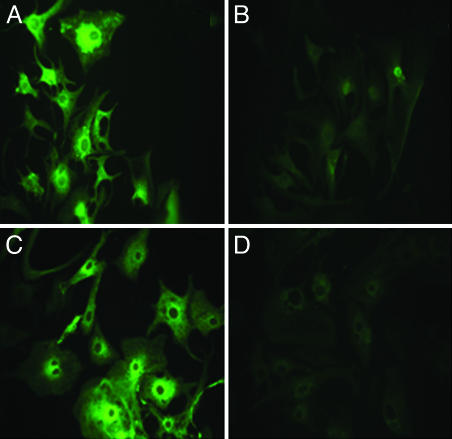
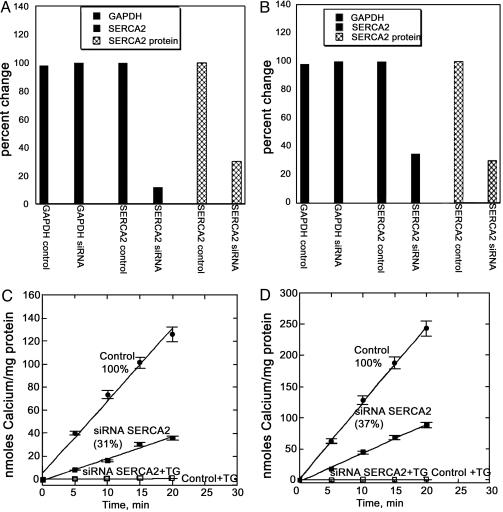
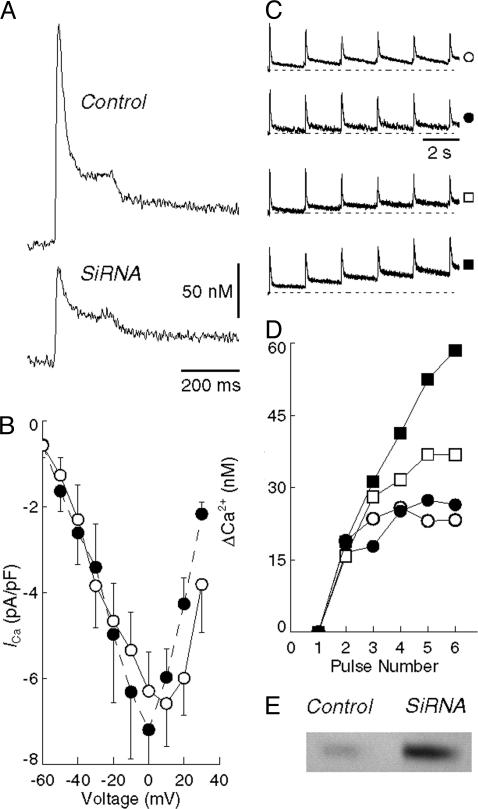
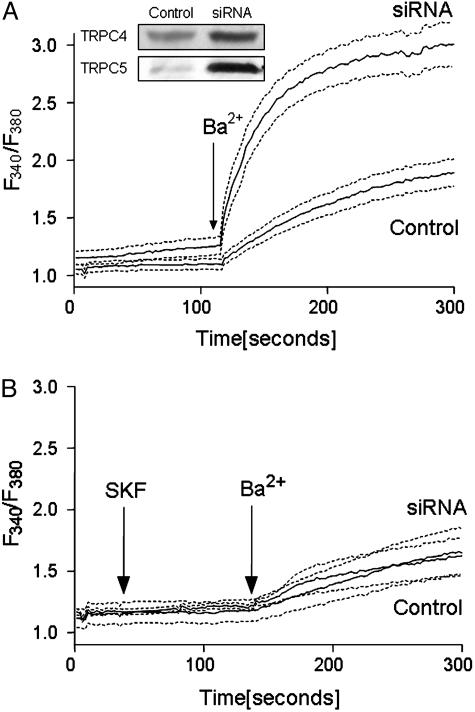
Similar articles
-
Phenylephrine hypertrophy, Ca2+-ATPase (SERCA2), and Ca2+ signaling in neonatal rat cardiac myocytes.Am J Physiol Cell Physiol. 2007 Jun;292(6):C2269-75. doi: 10.1152/ajpcell.00441.2006. Epub 2007 Feb 7. Am J Physiol Cell Physiol. 2007. PMID: 17287366
-
Cell-specific expression of SERCA, the exogenous Ca2+ transport ATPase, in cardiac myocytes.Am J Physiol Cell Physiol. 2004 Mar;286(3):C556-64. doi: 10.1152/ajpcell.00328.2003. Epub 2003 Oct 30. Am J Physiol Cell Physiol. 2004. PMID: 14592812
-
Co-ordinated regulation of the plasma membrane calcium pump and the sarco(endo)plasmic reticular calcium pump gene expression by Ca2+.Cell Calcium. 1997 Jun;21(6):399-408. doi: 10.1016/s0143-4160(97)90051-8. Cell Calcium. 1997. PMID: 9223676
-
Regulation of sarco(endo)plasmic reticulum Ca2+-ATPase and calsequestrin gene expression in the heart.Can J Physiol Pharmacol. 2012 Aug;90(8):1017-28. doi: 10.1139/y2012-057. Epub 2012 Jul 11. Can J Physiol Pharmacol. 2012. PMID: 22784385 Review.
-
SERCA in genesis of arrhythmias: what we already know and what is new?Anadolu Kardiyol Derg. 2007 Jul;7 Suppl 1:43-6. Anadolu Kardiyol Derg. 2007. PMID: 17584678 Review.
Cited by
-
Tissue-specific expression of TRP channel genes in the mouse and its variation in three different mouse strains.BMC Genomics. 2006 Jun 20;7:159. doi: 10.1186/1471-2164-7-159. BMC Genomics. 2006. PMID: 16787531 Free PMC article.
-
Molecular determinants of cardiac fibroblast electrical function and therapeutic implications for atrial fibrillation.Cardiovasc Res. 2011 Mar 1;89(4):744-53. doi: 10.1093/cvr/cvq329. Epub 2010 Oct 20. Cardiovasc Res. 2011. PMID: 20962103 Free PMC article. Review.
-
Roles of TRP channels in the development of cardiac hypertrophy.Naunyn Schmiedebergs Arch Pharmacol. 2008 Oct;378(4):395-406. doi: 10.1007/s00210-008-0321-8. Epub 2008 Jul 4. Naunyn Schmiedebergs Arch Pharmacol. 2008. PMID: 18600314 Review.
-
Regulation and rate limiting mechanisms of Ca2+ ATPase (SERCA2) expression in cardiac myocytes.Mol Cell Biochem. 2012 Feb;361(1-2):85-96. doi: 10.1007/s11010-011-1092-y. Epub 2011 Oct 1. Mol Cell Biochem. 2012. PMID: 21964539
-
Phospholamban as a crucial determinant of the inotropic response of human pluripotent stem cell-derived ventricular cardiomyocytes and engineered 3-dimensional tissue constructs.Circ Arrhythm Electrophysiol. 2015 Feb;8(1):193-202. doi: 10.1161/CIRCEP.114.002049. Epub 2014 Dec 10. Circ Arrhythm Electrophysiol. 2015. PMID: 25504561 Free PMC article.
References
-
- Carafoli, E., Santella, L., Branca, D. & Brini, M. (2001) Crit. Rev. Biochem. Mol. Biol. 36, 107–260. - PubMed
-
- Berridge, M. J., Bootman, M. D. & Roderick, H. L. (2003) Nat. Rev. Mol. Cell Biol. 4, 517–529. - PubMed
-
- Eisner, D. A., Choi, H. S., Diaz, M. E., O'Neill, S. C. & Trafford, A. W. (2000) Circ. Res. 87, 1087–1094. - PubMed
-
- Bers, D. M. (2002) Nature 415, 198–205. - PubMed
-
- MacLennan, D. H. & Kranias, E. G. (2003) Nat. Rev. Mol. Cell Biol. 4, 566–577. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
