

A physiological connection between tmRNA and peptidyl-tRNA hydrolase functions in Escherichia coli
- PMID: 15547251
- PMCID: PMC534616
- DOI: 10.1093/nar/gkh924
A physiological connection between tmRNA and peptidyl-tRNA hydrolase functions in Escherichia coli
Abstract
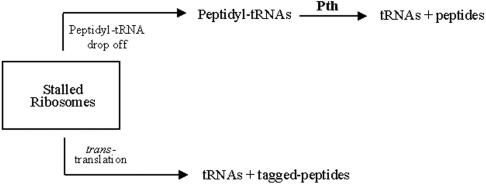
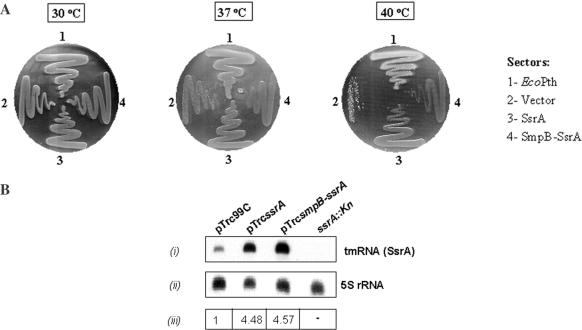
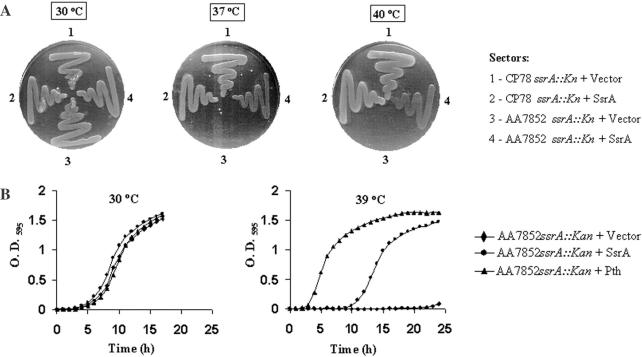
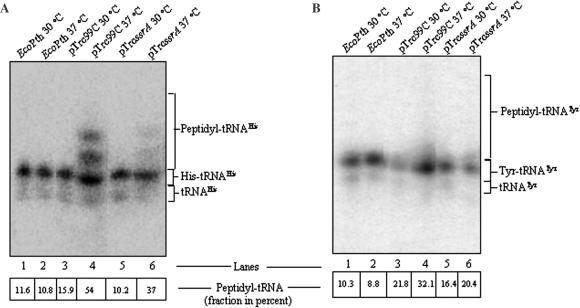
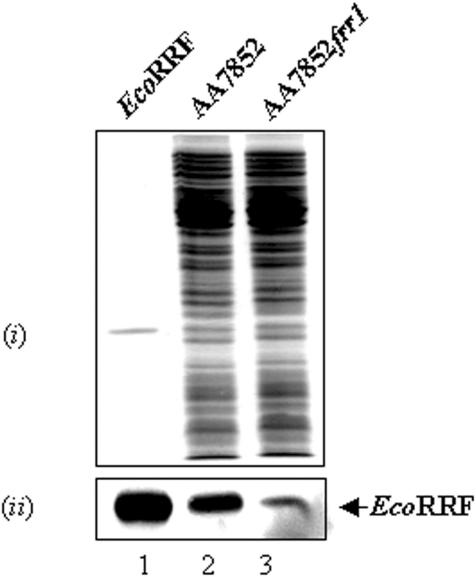
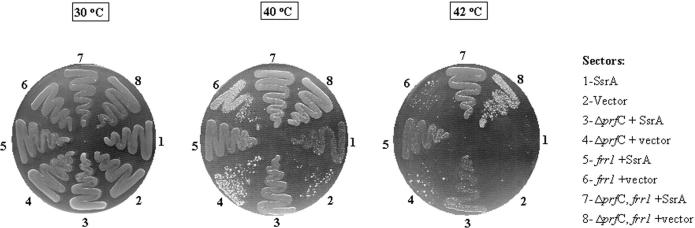
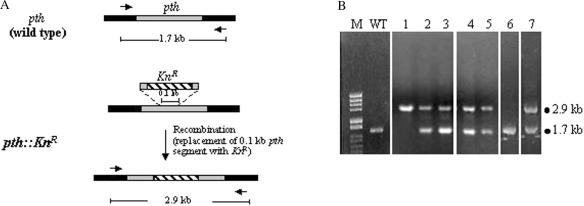
The bacterial ssrA gene codes for a dual function RNA, tmRNA, which possesses tRNA-like and mRNA-like regions. The tmRNA appends an oligopeptide tag to the polypeptide on the P-site tRNA by a trans-translation process that rescues ribosomes stalled on the mRNAs and targets the aberrant protein for degradation. In cells, processing of the stalled ribosomes is also pioneered by drop-off of peptidyl-tRNAs. The ester bond linking the peptide to tRNA is hydrolyzed by peptidyl-tRNA hydrolase (Pth), an essential enzyme, which releases the tRNA and the aberrant peptide. As the trans-translation mechanism utilizes the peptidyl-transferase activity of the stalled ribosomes to free the tRNA (as opposed to peptidyl-tRNA drop-off), the need for Pth to recycle such tRNAs is bypassed. Thus, we hypothesized that tmRNA may rescue a defect in Pth. Here, we show that overexpression of tmRNA rescues the temperature-sensitive phenotype of Escherichia coli (pth(ts)). Conversely, a null mutation in ssrA enhances the temperature-sensitive phenotype of the pth(ts) strain. Consistent with our hypothesis, overexpression of tmRNA results in decreased accumulation of peptidyl-tRNA in E.coli. Furthermore, overproduction of tmRNA in E.coli strains deficient in ribosome recycling factor and/or lacking the release factor 3 enhances the rescue of pth(ts) strains. We discuss the physiological relevance of these observations to highlight a major role of tmRNA in decreasing cellular peptidyl-tRNA load.
Figures
Similar articles
-
Ribosome recycling factor and release factor 3 action promotes TnaC-peptidyl-tRNA Dropoff and relieves ribosome stalling during tryptophan induction of tna operon expression in Escherichia coli.J Bacteriol. 2007 Apr;189(8):3147-55. doi: 10.1128/JB.01868-06. Epub 2007 Feb 9. J Bacteriol. 2007. PMID: 17293419 Free PMC article.
-
Protein synthesis factors (RF1, RF2, RF3, RRF, and tmRNA) and peptidyl-tRNA hydrolase rescue stalled ribosomes at sense codons.J Mol Biol. 2012 Apr 13;417(5):425-39. doi: 10.1016/j.jmb.2012.02.008. Epub 2012 Feb 9. J Mol Biol. 2012. PMID: 22326347
-
Analysis of nonstop mRNA translation in the absence of tmRNA in Escherichia coli.Genes Cells. 2009 Jun;14(6):739-49. doi: 10.1111/j.1365-2443.2009.01304.x. Epub 2009 May 14. Genes Cells. 2009. PMID: 19469882
-
A salvage pathway for protein structures: tmRNA and trans-translation.Annu Rev Microbiol. 2003;57:101-23. doi: 10.1146/annurev.micro.57.030502.090945. Epub 2003 May 1. Annu Rev Microbiol. 2003. PMID: 12730326 Review.
-
Trans-translation: the tmRNA-mediated surveillance mechanism for ribosome rescue, directed protein degradation, and nonstop mRNA decay.Biochemistry. 2007 Apr 24;46(16):4681-93. doi: 10.1021/bi6026055. Epub 2007 Mar 31. Biochemistry. 2007. PMID: 17397189 Review.
Cited by
-
Ribosome Rescue Pathways in Bacteria.Front Microbiol. 2021 Mar 18;12:652980. doi: 10.3389/fmicb.2021.652980. eCollection 2021. Front Microbiol. 2021. PMID: 33815344 Free PMC article. Review.
-
Laboratory evolution and multi-platform genome re-sequencing of the cellulolytic actinobacterium Thermobifida fusca.J Biol Chem. 2011 Nov 18;286(46):39958-66. doi: 10.1074/jbc.M111.239616. Epub 2011 Sep 13. J Biol Chem. 2011. PMID: 21914801 Free PMC article.
-
Complementary roles of initiation factor 1 and ribosome recycling factor in 70S ribosome splitting.EMBO J. 2008 Jun 18;27(12):1706-17. doi: 10.1038/emboj.2008.99. Epub 2008 May 22. EMBO J. 2008. PMID: 18497739 Free PMC article.
-
The tmRNA ribosome-rescue system.Adv Protein Chem Struct Biol. 2012;86:151-91. doi: 10.1016/B978-0-12-386497-0.00005-0. Adv Protein Chem Struct Biol. 2012. PMID: 22243584 Free PMC article. Review.
-
Synthesis of a Thiazole Library via an Iridium-Catalyzed Sulfur Ylide Insertion Reaction.Org Lett. 2022 Nov 4;24(43):7924-7927. doi: 10.1021/acs.orglett.2c02996. Epub 2022 Oct 20. Org Lett. 2022. PMID: 36265082 Free PMC article.
References
-
- Manley J.L. (1978) Synthesis and degradation of termination and premature-termination fragments of beta-galactosidase in vitro and in vivo. J. Mol. Biol., 125, 407–432. - PubMed
-
- Jorgensen F. and Kurland,C.G. (1990) Processivity errors of gene expression in Escherichia coli. J. Mol. Biol., 215, 511–521. - PubMed
-
- Menninger J.R. (1976) Peptidyl transfer RNA dissociates during protein synthesis from ribosomes of Escherichia coli. J. Biol. Chem., 251, 3392–3398. - PubMed
-
- Menninger J.R. (1978) The accumulation as peptidyl-transfer RNA of isoaccepting transfer RNA families in Escherichia coli with temperature-sensitive peptidyl-transfer RNA hydrolase. J. Biol. Chem., 253, 6808–6813. - PubMed
-
- Caplan A.B. and Menninger,J.R. (1979) Tests of the ribosomal editing hypothesis: amino acid starvation differentially enhances the dissociation of peptidyl-tRNA from the ribosome. J. Mol. Biol., 134, 621–637. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
