

In vivo activity of enzymatic and regulatory components of the phosphoenolpyruvate:sugar phosphotransferase system in Mycoplasma pneumoniae
- PMID: 15547265
- PMCID: PMC529091
- DOI: 10.1128/JB.186.23.7936-7943.2004
In vivo activity of enzymatic and regulatory components of the phosphoenolpyruvate:sugar phosphotransferase system in Mycoplasma pneumoniae
Abstract
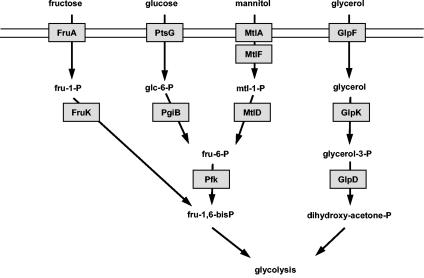
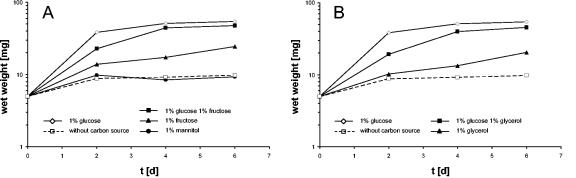
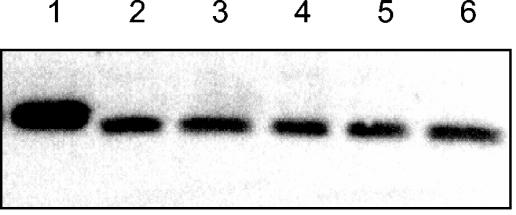
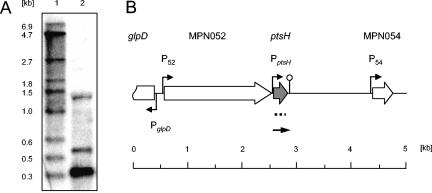
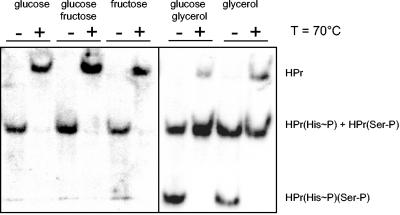
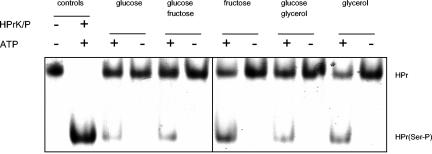
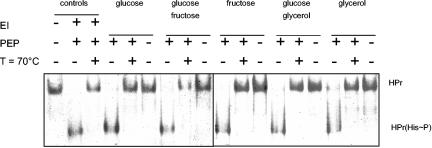
Mycoplasma pneumoniae is a pathogenic bacterium that is highly adapted to life on mucosal surfaces. This adaptation is reflected by the very compact genome and the small number of regulatory proteins. However, M. pneumoniae possesses the HPr kinase/phosphorylase (HPrK/P), the key regulator of carbon metabolism in the Firmicutes. In contrast to the enzymes of other bacteria, the HPrK/P of M. pneumoniae is already active at very low ATP concentrations, suggesting a different mode of regulation. In this work, we studied the ability of M. pneumoniae to utilize different carbohydrates and their effects on the activity of the different phosphotransferase system (PTS) components. Glucose served as the best carbon source, with a generation time of about 30 h. Fructose and glycerol were also used but at lower rates and with lower yields. In contrast, M. pneumoniae is unable to use mannitol even though the bacterium is apparently equipped with all the genes required for mannitol catabolism. This observation is probably a reflection of the continuing and ongoing reduction of the M. pneumoniae genome. The general enzymatic and regulatory components of the PTS, i.e., enzyme I, HPr, and HPrK/P, were present under all growth conditions tested in this study. However, HPrK/P activity is strongly increased if the medium contains glycerol. Thus, the control of HPrK/P in vivo differs strongly between M. pneumoniae and the other Firmicutes. This difference may relate to the specific conditions on lipid-rich cell surfaces.
Figures
References
-
- Allen, G. S., K. Steinhauer, W. Hillen, J. Stülke, and R. G. Brennan. 2003. Crystal structure of HPr kinase/phosphatase from Mycoplasma pneumoniae. J. Mol. Biol. 326:1203-1217. - PubMed
-
- Bradford, M. M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72:248-254. - PubMed
-
- Dandekar, T., M. Huynen, J. T. Regula, B. Ueberle, C. U. Zimmermann, M. A. Andrade, T. Doerks, L. Sánchez-Pulido, B. Snel, M. Suyama, Y. P. Yuan, R. Herrmann, and P. Bork. 2000. Re-annotating the Mycoplasma pneumoniae genome sequence: adding value, function and reading frames. Nucleic Acids Res. 28:3278-3288. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
