

The Caenorhabditis elegans Aurora B kinase AIR-2 phosphorylates and is required for the localization of a BimC kinesin to meiotic and mitotic spindles
- PMID: 15548597
- PMCID: PMC545908
- DOI: 10.1091/mbc.e04-08-0682
The Caenorhabditis elegans Aurora B kinase AIR-2 phosphorylates and is required for the localization of a BimC kinesin to meiotic and mitotic spindles
Abstract
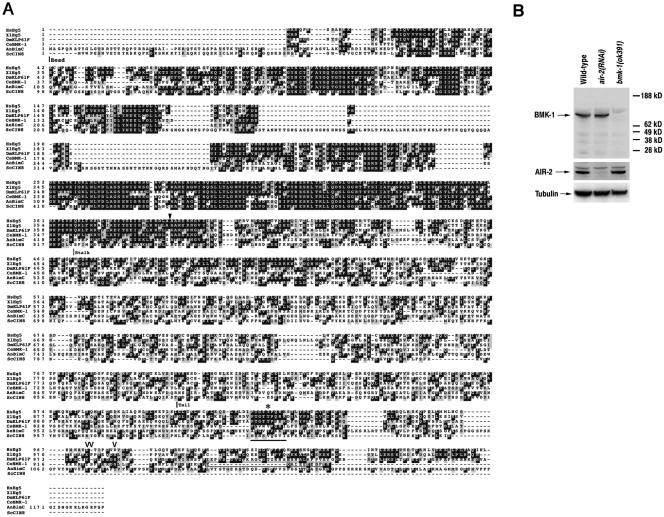
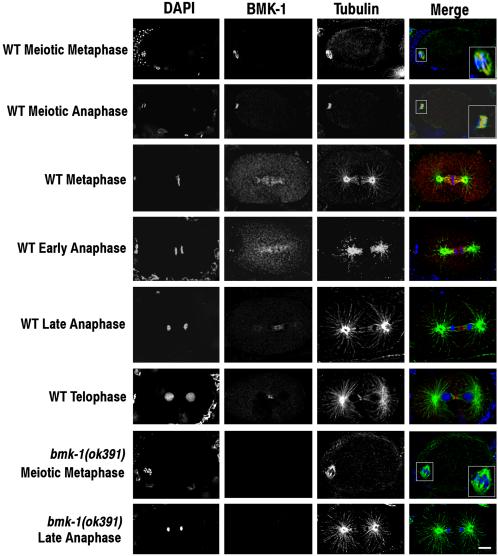
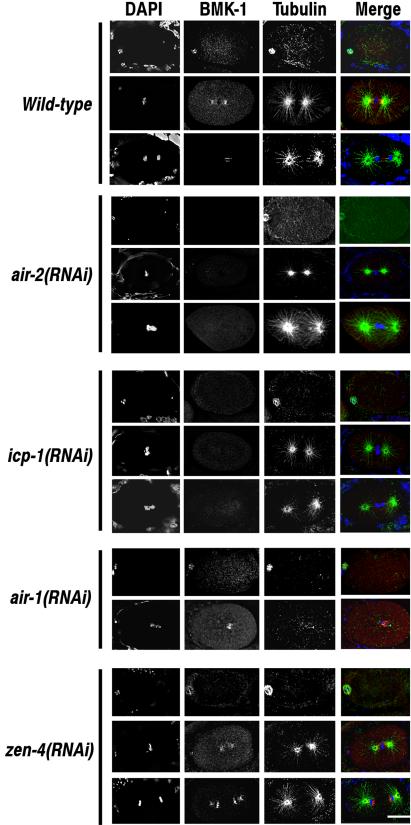
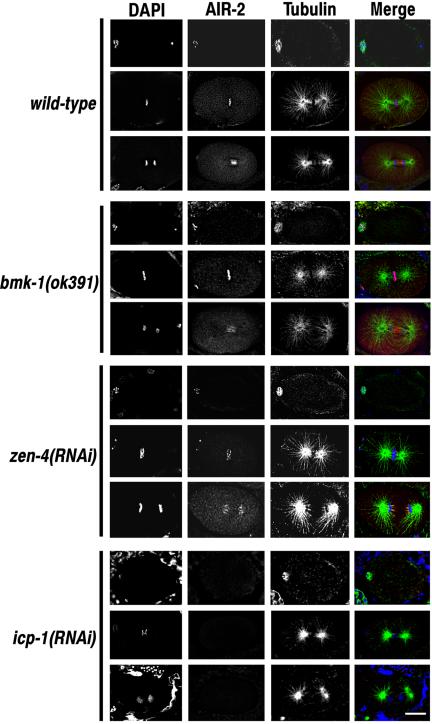
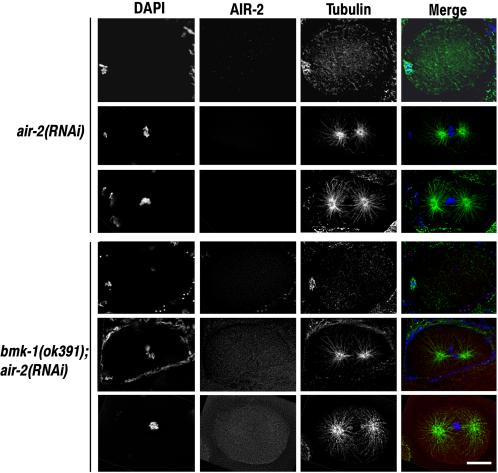
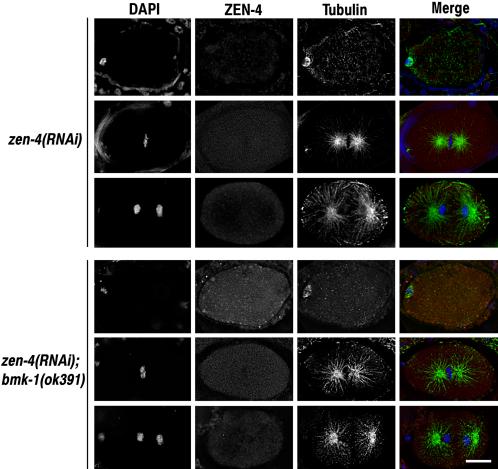
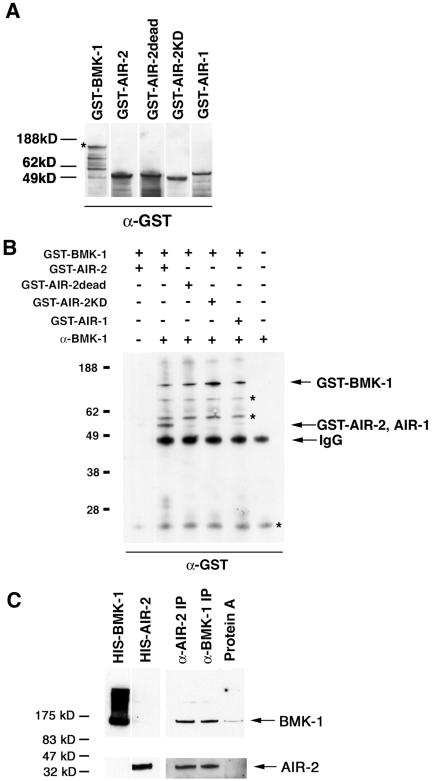
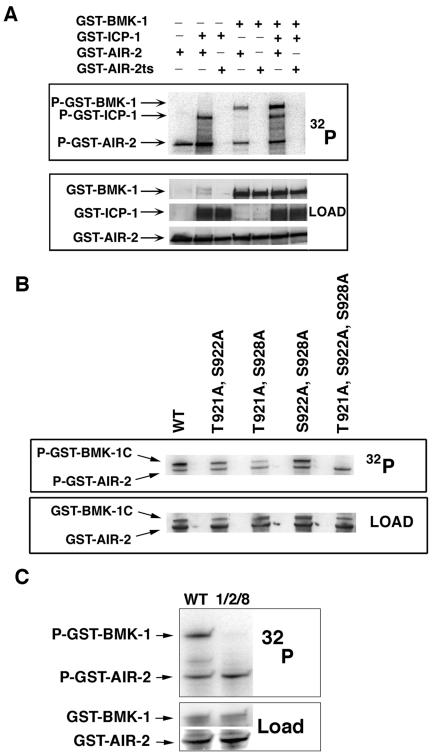
BimC kinesins are required for mitotic spindle assembly in a variety of organisms. These proteins are localized to centrosomes, spindle microtubules, and the spindle midzone. We have previously shown that the Caenorhabditis elegans Aurora B kinase AIR-2 is required for the localization of the ZEN-4 kinesin protein to midzone microtubules. To determine whether the association of BimC kinesins with spindle microtubules is also dependent on AIR-2, we examined the expression pattern of BMK-1, a C. elegans BimC kinesin, in wild-type and AIR-2-deficient embryos. BMK-1 is highly expressed in the hermaphrodite gonad and is localized to meiotic spindle microtubules in the newly fertilized embryo. In mitotic embryos, BMK-1 is associated with spindle microtubules from prophase through anaphase and is concentrated at the spindle midzone during anaphase and telophase. In the absence of AIR-2, BMK-1 localization to meiotic and mitotic spindles is greatly reduced. This is not a consequence of loss of ZEN-4 localization because BMK-1 is appropriately localized in ZEN-4-deficient embryos. Furthermore, AIR-2 and BMK-1 directly interact with one another and the C-terminal tail domain of BMK-1 is specifically phosphorylated by AIR-2 in vitro. Together with our previous data, these results suggest that at least one function of the Aurora B kinases is to recruit spindle-associated motor proteins to their sites of action.
Figures
References
-
- Adams, R. R., Carmena, M., and Earnshaw, W. C. (2001). Chromosomal passengers and the (aurora) ABCs of mitosis. Trends Cell Biol. 11, 49-54. - PubMed
-
- Bischoff, J. R., and Plowman, G. D. (1999). The Aurora/Ipl1p kinase family: regulators of chromosome segregation and cytokinesis. Trends Cell Biol. 9, 454-459. - PubMed
-
- Blangy, A., Lane, H. A., d'Herin, P., Harper, M., Kress, M., and Nigg, E. A. (1995). Phosphorylation by p34cdc2 regulates spindle association of human Eg5, a kinesin-related motor essential for bipolar spindle formation in vivo. Cell 83, 1159-1169. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
