

The Arabidopsis peroxisomal targeting signal type 2 receptor PEX7 is necessary for peroxisome function and dependent on PEX5
- PMID: 15548601
- PMCID: PMC545895
- DOI: 10.1091/mbc.e04-05-0422
The Arabidopsis peroxisomal targeting signal type 2 receptor PEX7 is necessary for peroxisome function and dependent on PEX5
Abstract
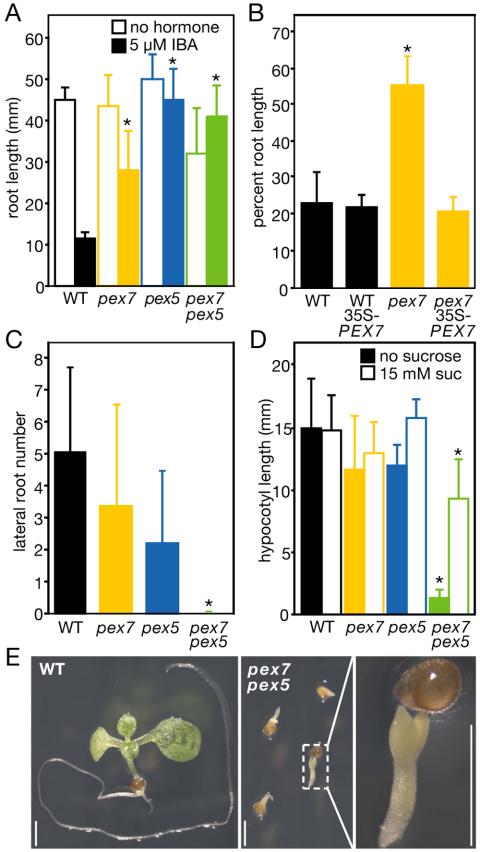
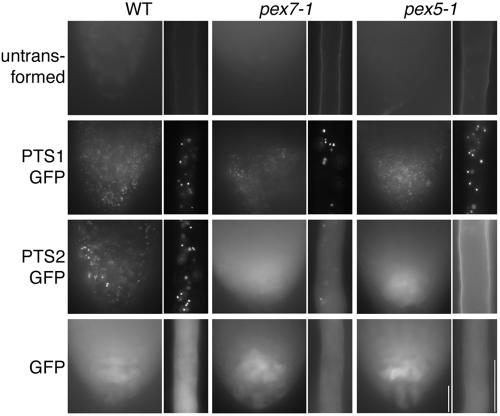
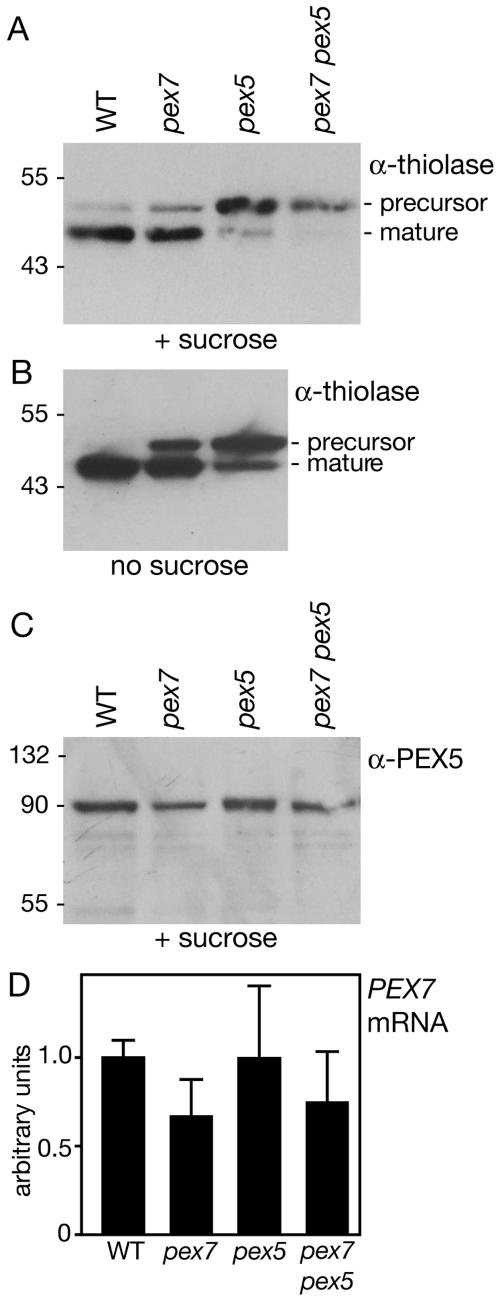
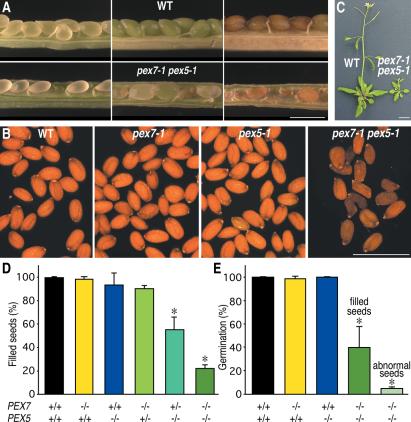
Plant peroxisomal proteins catalyze key metabolic reactions. Several peroxisome biogenesis PEROXIN (PEX) genes encode proteins acting in the import of targeted proteins necessary for these processes into the peroxisomal matrix. Most peroxisomal matrix proteins bear characterized Peroxisomal Targeting Signals (PTS1 or PTS2), which are bound by the receptors PEX5 or PEX7, respectively, for import into peroxisomes. Here we describe the isolation and characterization of an Arabidopsis peroxin mutant, pex7-1, which displays peroxisome-defective phenotypes including reduced PTS2 protein import. We also demonstrate that the pex5-1 PTS1 receptor mutant, which contains a lesion in a domain conserved among PEX7-binding proteins from various organisms, is defective not in PTS1 protein import, but rather in PTS2 protein import. Combining these mutations in a pex7-1 pex5-1 double mutant abolishes detectable PTS2 protein import and yields seedlings that are entirely sucrose-dependent for establishment, suggesting a severe block in peroxisomal fatty acid beta-oxidation. Adult pex7-1 pex5-1 plants have reduced stature and bear abnormally shaped seeds, few of which are viable. The pex7-1 pex5-1 seedlings that germinate have dramatically fewer lateral roots and often display fused cotyledons, phenotypes associated with reduced auxin response. Thus PTS2-directed peroxisomal import is necessary for normal embryonic development, seedling establishment, and vegetative growth.
Figures



References
-
- Aida, M., Vernoux, T., Furutani, M., Traas, J., and Tasaka, M. (2002). Roles of PIN-FORMED1 and MONOPTEROS in pattern formation of the apical region of the Arabidopsis embryo. Development 129, 3965-3974. - PubMed
-
- Albertini, M., Rehling, P., Erdmann, R., Girzalsky, W., Kiel, J.A.K.W., Veenhuis, M., and Kunau, W. H. (1997). Pex14p, a peroxisomal membrane protein binding both receptors of the two PTS-dependent import pathways. Cell 89, 83-92. - PubMed
-
- Alonso, J. M. et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301, 653-657. - PubMed
-
- Asamizu, E., Nakamura, Y., Sato, S., and Tabata, S. (2000). A large scale analysis of cDNA in Arabidopsis thaliana: generation of 12,028 non-redundant expressed sequence tags from normalized and size-selected cDNA libraries. DNA Res. 7, 175-180. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
