

Acetylcholine dynamically controls spatial integration in marmoset primary visual cortex
- PMID: 15548624
- PMCID: PMC1891447
- DOI: 10.1152/jn.00911.2004
Acetylcholine dynamically controls spatial integration in marmoset primary visual cortex
Abstract
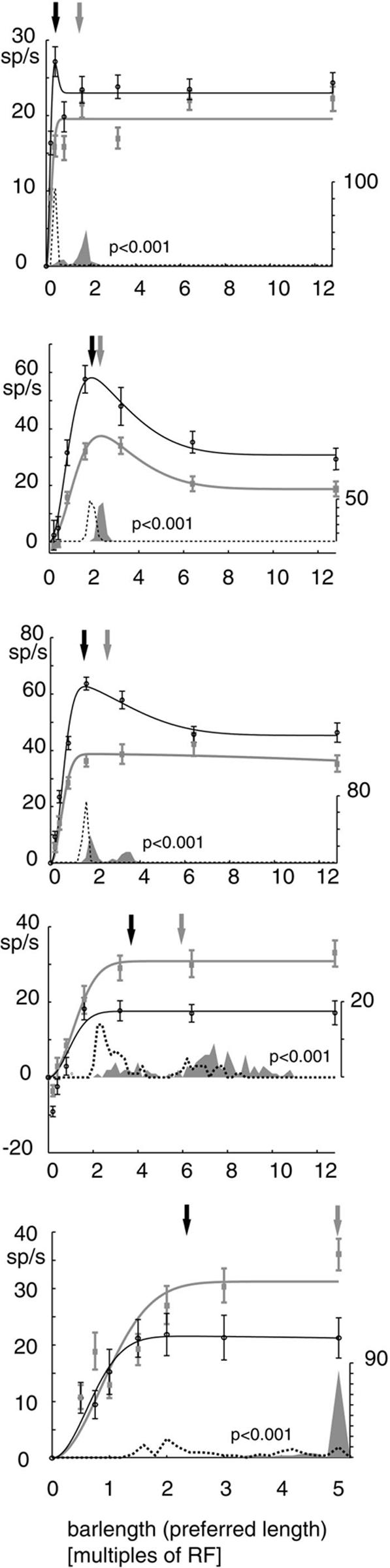
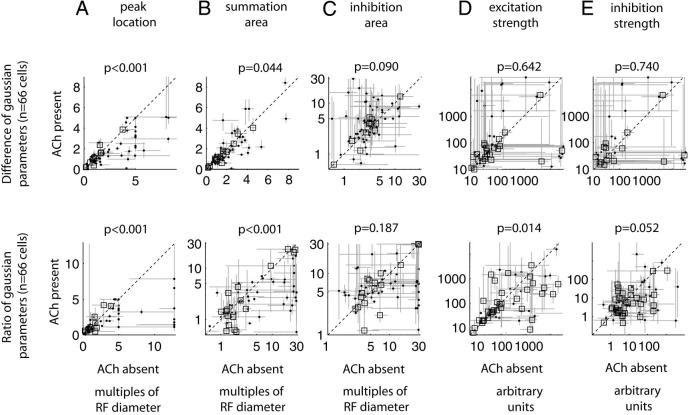
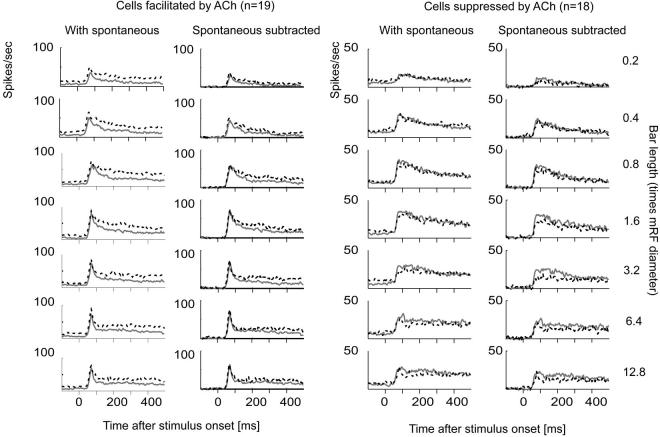
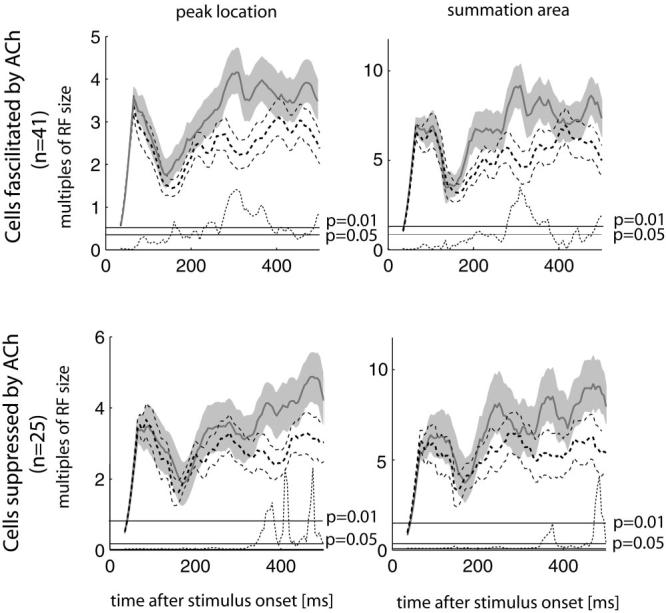
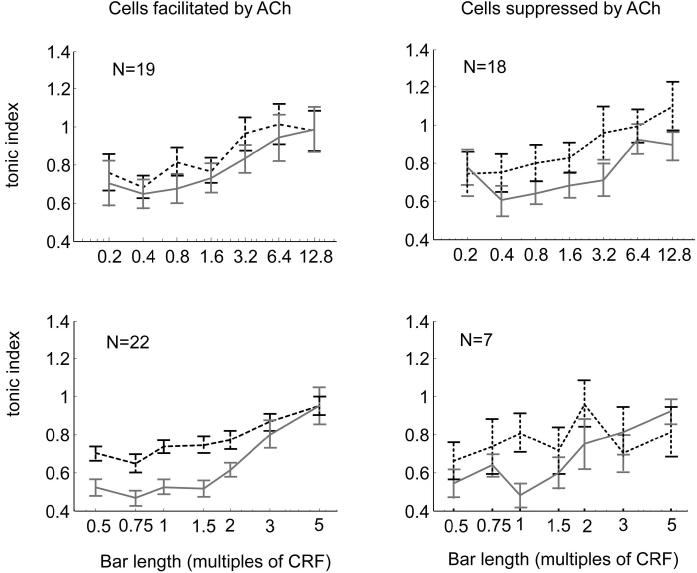
Recent in vitro studies have shown that acetylcholine (ACh) selectively reduces the efficacy of lateral cortical connections via a muscarinic mechanism, while boosting the efficacy of thalamocortical/feed-forward connections via a nicotinic mechanism. This suggests that high levels of ACh should reduce center-surround interactions of neurons in primary visual cortex, making cells more reliant on feed-forward information. In line with this hypothesis, we show that local iontophoretic application of ACh in primate primary visual cortex reduced the extent of spatial integration, assessed by recording a neurons' length tuning. When ACh was externally applied, neurons' preferred length shifted toward shorter bars, showing reduced impact of the extra-classical receptive field. We fitted a difference and a ratio of Gaussian model to these data to determine the underlying mechanisms of this dynamic change of spatial integration. These models assume overlapping summation and suppression areas with different widths and gains to be responsible for spatial integration and size tuning. ACh significantly reduced the extent of the summation area, but had no significant effect on the extent of the suppression area. In line with previous studies, we also show that applying ACh enhanced the response in the majority of cells, especially in the later (sustained) part of the response. These findings are similar to effects of attention on neuronal activity. The natural release of ACh is strongly linked with states of arousal and attention. Our results may therefore be relevant to the neurobiological mechanism of attention.
Figures
Similar articles
-
Cholinergic modulation of response properties and orientation tuning of neurons in primary visual cortex of anaesthetized Marmoset monkeys.Eur J Neurosci. 2006 Jul;24(1):314-28. doi: 10.1111/j.1460-9568.2006.04882.x. Eur J Neurosci. 2006. PMID: 16882027 Free PMC article.
-
Spatial integration and its moderation by attention and acetylcholine.Front Biosci. 2008 May 1;13:3742-59. doi: 10.2741/2963. Front Biosci. 2008. PMID: 18508469
-
Modulation-specific and laminar-dependent effects of acetylcholine on visual responses in the rat primary visual cortex.PLoS One. 2013 Jul 2;8(7):e68430. doi: 10.1371/journal.pone.0068430. Print 2013. PLoS One. 2013. PMID: 23844199 Free PMC article.
-
Contribution of feedforward, lateral and feedback connections to the classical receptive field center and extra-classical receptive field surround of primate V1 neurons.Prog Brain Res. 2006;154:93-120. doi: 10.1016/S0079-6123(06)54005-1. Prog Brain Res. 2006. PMID: 17010705 Review.
-
The contribution of vertical and horizontal connections to the receptive field center and surround in V1.Neural Netw. 2004 Jun-Jul;17(5-6):681-93. doi: 10.1016/j.neunet.2004.05.002. Neural Netw. 2004. PMID: 15288892 Review.
Cited by
-
Neurochemistry of Visual Attention.Front Neurosci. 2021 May 5;15:643597. doi: 10.3389/fnins.2021.643597. eCollection 2021. Front Neurosci. 2021. PMID: 34025339 Free PMC article. Review.
-
Contribution of cholinergic and GABAergic mechanisms to direction tuning, discriminability, response reliability, and neuronal rate correlations in macaque middle temporal area.J Neurosci. 2012 Nov 21;32(47):16602-15. doi: 10.1523/JNEUROSCI.0554-12.2012. J Neurosci. 2012. PMID: 23175816 Free PMC article.
-
An acetylcholine-activated microcircuit drives temporal dynamics of cortical activity.Nat Neurosci. 2015 Jun;18(6):892-902. doi: 10.1038/nn.4002. Epub 2015 Apr 27. Nat Neurosci. 2015. PMID: 25915477 Free PMC article.
-
Posterior parietal cortex dynamically ranks topographic signals via cholinergic influence.Front Integr Neurosci. 2012 Jun 14;6:32. doi: 10.3389/fnint.2012.00032. eCollection 2012. Front Integr Neurosci. 2012. PMID: 22712008 Free PMC article.
-
Cholinergic Modulation Promotes Attentional Modulation in Primary Visual Cortex- A Modeling Study.Sci Rep. 2019 Dec 27;9(1):20186. doi: 10.1038/s41598-019-56608-3. Sci Rep. 2019. PMID: 31882838 Free PMC article.
References
-
- Abbott LF, Varela JA, Sen K, Nelson SB. Synaptic depression and cortical gain control. Science. 1997;275:220–224. - PubMed
-
- Albright TD, Stoner GR. Contextual influences on visual processing. Annu Rev Neurosci. 2002;25:339–379. - PubMed
-
- Angelucci A, Levitt JB, Lund JS. Anatomical origins of the classical receptive field and modulatory surround field of single neurons in macaque visual cortical area V1. Prog Brain Res. 2002;136:373–388. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources