

Functional compensation of P/Q by N-type channels blocks short-term plasticity at the calyx of Held presynaptic terminal
- PMID: 15548652
- PMCID: PMC6730293
- DOI: 10.1523/JNEUROSCI.2104-04.2004
Functional compensation of P/Q by N-type channels blocks short-term plasticity at the calyx of Held presynaptic terminal
Abstract
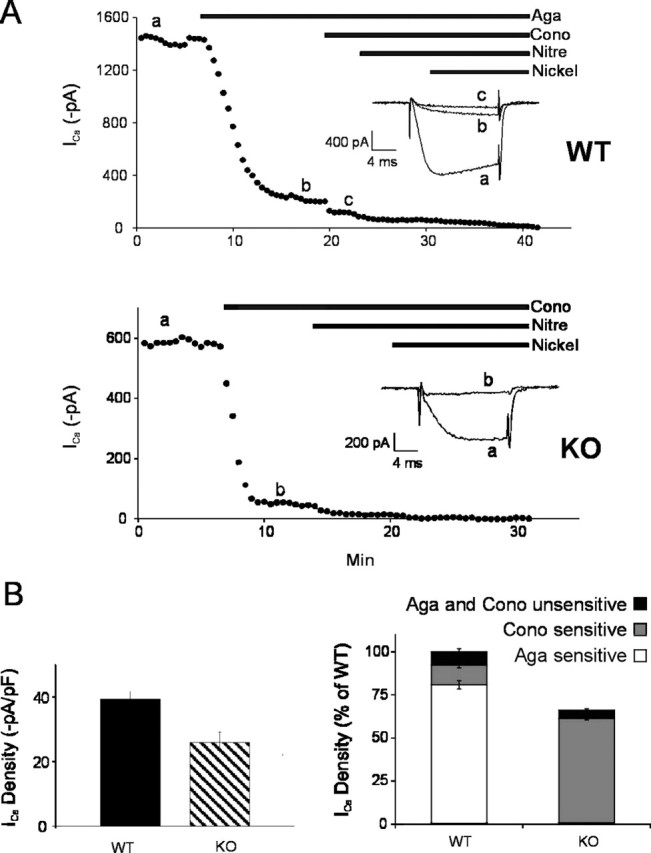
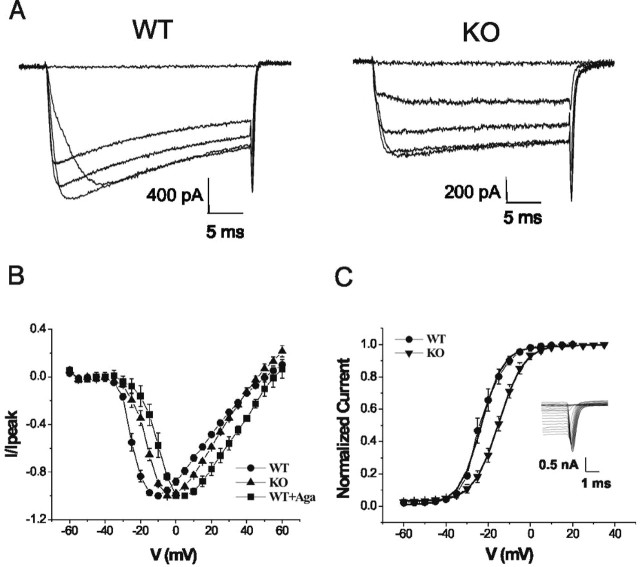
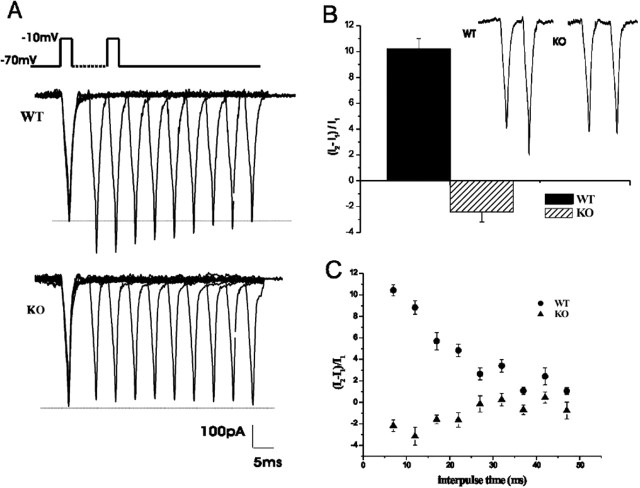
Calcium channels of the P/Q subtype mediate transmitter release at the neuromuscular junction and at many central synapses, such as the calyx of Held. Transgenic mice in which alpha1A channels are ablated provide a powerful tool with which to test compensatory mechanisms at the synapse and to explore mechanisms of presynaptic regulation associated with expression of P/Q channels. Using the calyx of Held preparation from the knock-out (KO) mice, we show here that N-type channels functionally compensate for the absence of P/Q subunits at the calyx and evoke giant synaptic currents [approximately two-thirds of the magnitude of wild-type (WT) responses]. However, although evoked paired-pulse facilitation is prominent in WT, this facilitation is greatly diminished in the KO. In addition, direct recording of presynaptic calcium currents revealed that the major functional difference was the absence of calcium-dependent facilitation at the calyx in the P/Q KO animals. We conclude that one physiological function of P/Q channels is to provide additional facilitatory drive, so contributing to maintenance of transmission as vesicles are depleted during high throughput synaptic transmission.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials