

The role of palmitoylation in functional expression of nicotinic alpha7 receptors
- PMID: 15548665
- PMCID: PMC6730317
- DOI: 10.1523/JNEUROSCI.3315-04.2004
The role of palmitoylation in functional expression of nicotinic alpha7 receptors
Abstract
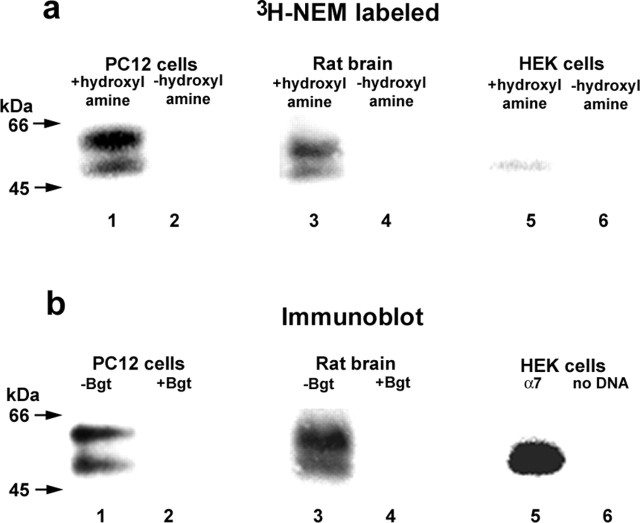
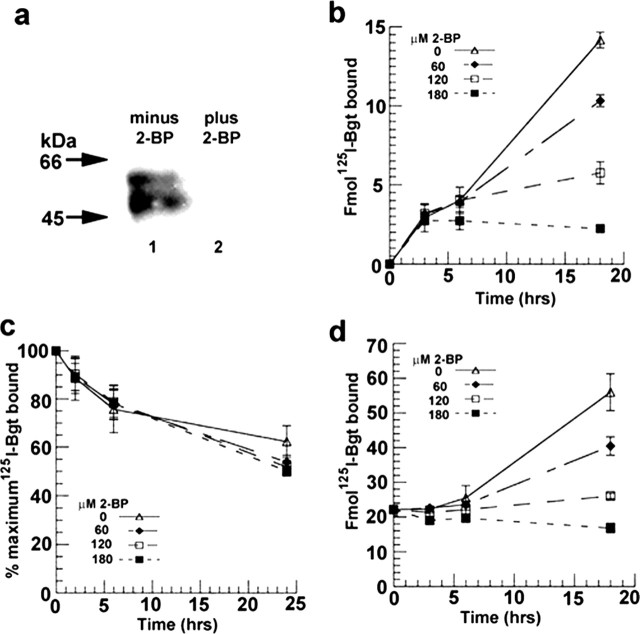
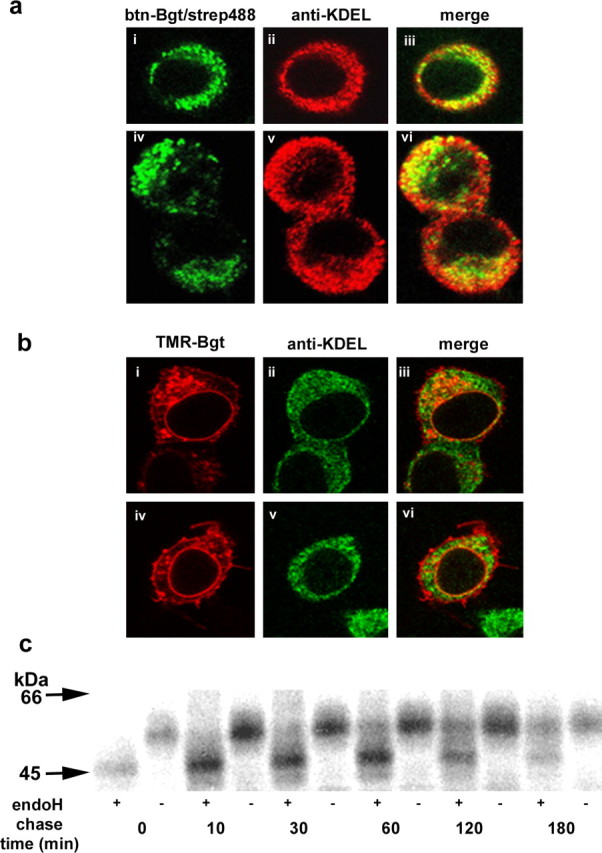
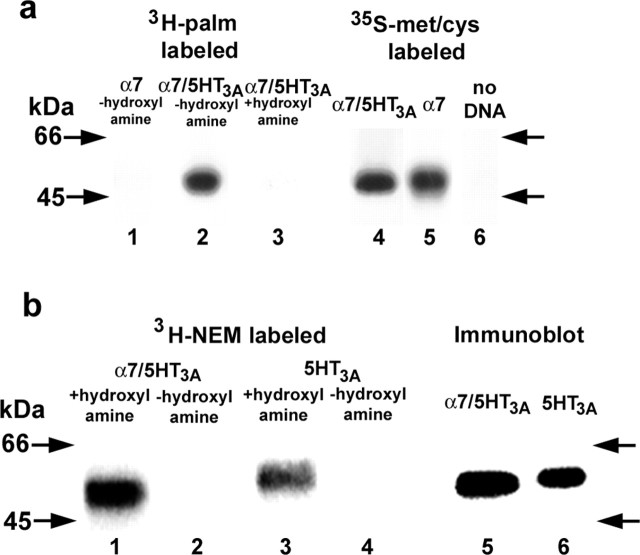
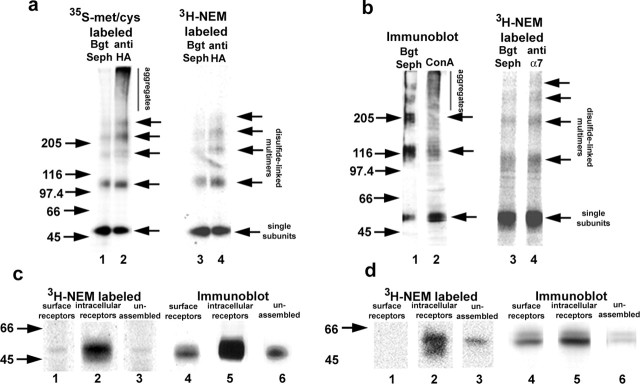
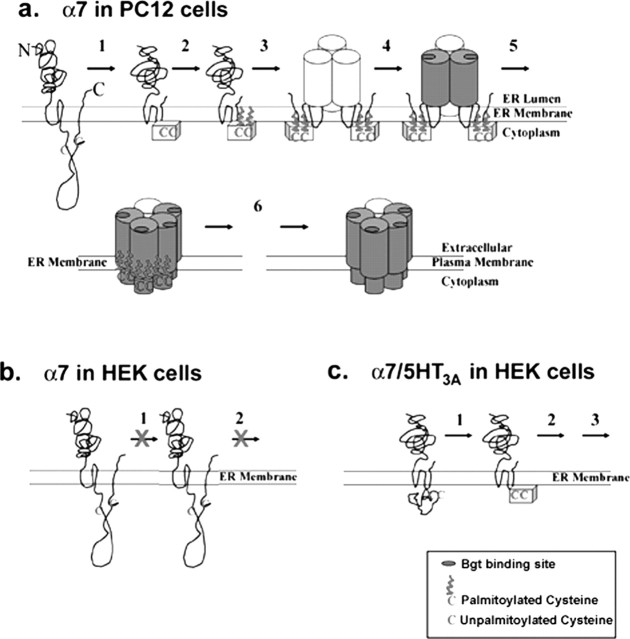
Neuronal alpha-bungarotoxin receptors (BgtRs) are nicotinic receptors that require as yet unidentified post-translational modifications to achieve functional expression. In this study, we examined the role of protein palmitoylation in BgtR expression. BgtR alpha7 subunits are highly palmitoylated in neurons from brain and other cells capable of BgtR expression, such as pheochromocytoma 12 (PC12) cells. In PC12 cells, alpha7 subunits are palmitoylated with a stoichiometry of approximately one palmitate per subunit, and inhibition of palmitoylation blocks BgtR expression. In cells incapable of BgtR expression, such as human embryonic kidney cells, alpha7 subunits are not significantly palmitoylated. However, in these same cells, chimeric subunits with the N-terminal half of alpha7 fused to the C-terminal half of serotonin-3A receptor (alpha7/5-HT3A) subunits form functional BgtRs that are palmitoylated to an extent similar to that of BgtRalpha7 subunits in PC12 cells. Palmitoylation of PC12 and alpha7/5-HT3A BgtRs occurred during assembly in the endoplasmic reticulum (ER). In conclusion, our data indicate a function for protein palmitoylation in which palmitoylation of assembling alpha7 subunits in the ER has a role in the formation of functional BgtRs.
Figures
References
-
- Barry MF, Ziff EB (2002) Receptor trafficking and the plasticity of excitatory synapses. Curr Opin Neurobiol 12: 279-286. - PubMed
-
- Bijlmakers MJ, Marsh M (2003) The on-off story of protein palmitoylation. Trends Cell Biol 13: 32-42. - PubMed
-
- Bizzozero OA, Malkoski SP, Mobarak C, Bixler HA, Evans JE (2002) Mass-spectrophotometric analysis of myelin proteolipids reveals new features of this family of palmitoylated membrane proteins. J Neurochem 81: 636-645. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources