

RAR1 positively controls steady state levels of barley MLA resistance proteins and enables sufficient MLA6 accumulation for effective resistance
- PMID: 15548741
- PMCID: PMC535887
- DOI: 10.1105/tpc.104.026682
RAR1 positively controls steady state levels of barley MLA resistance proteins and enables sufficient MLA6 accumulation for effective resistance
Abstract
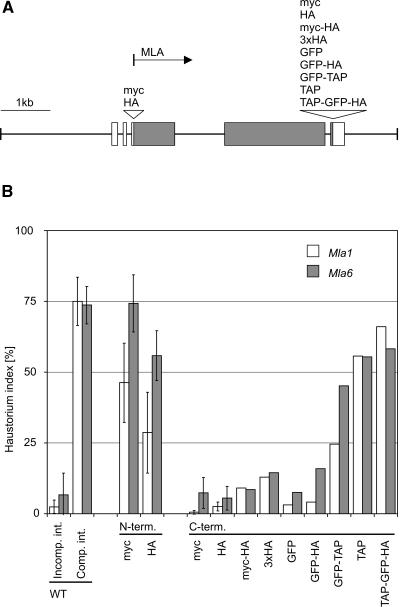
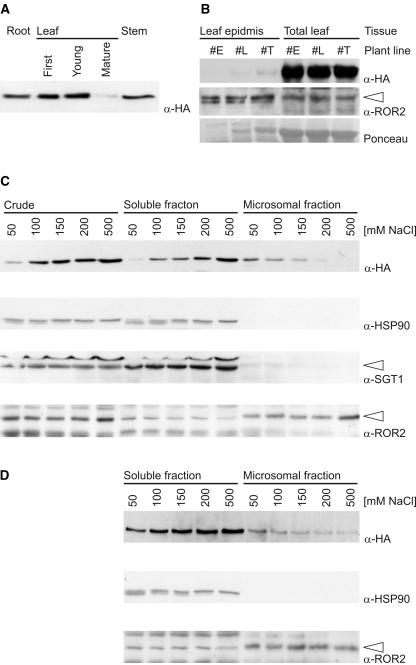
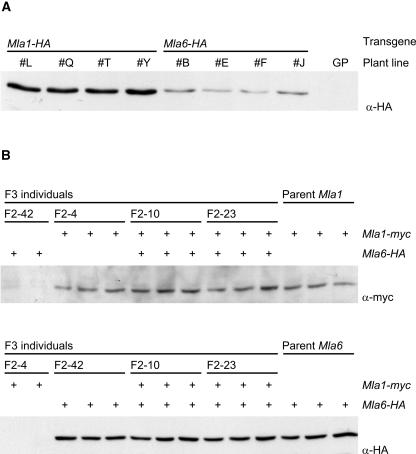
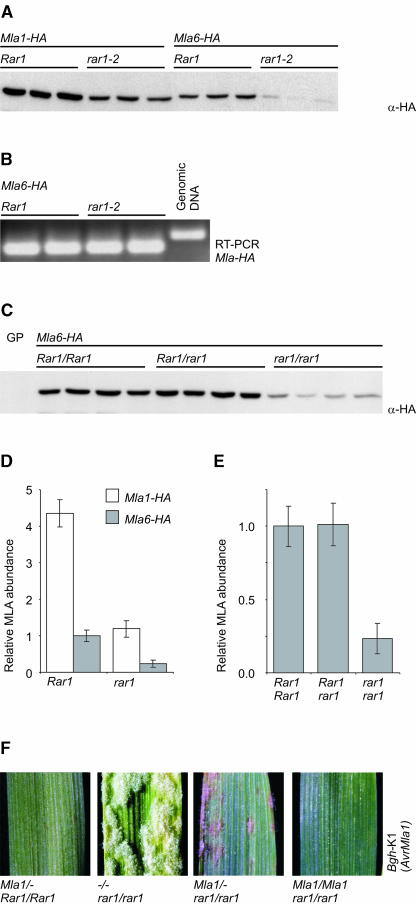
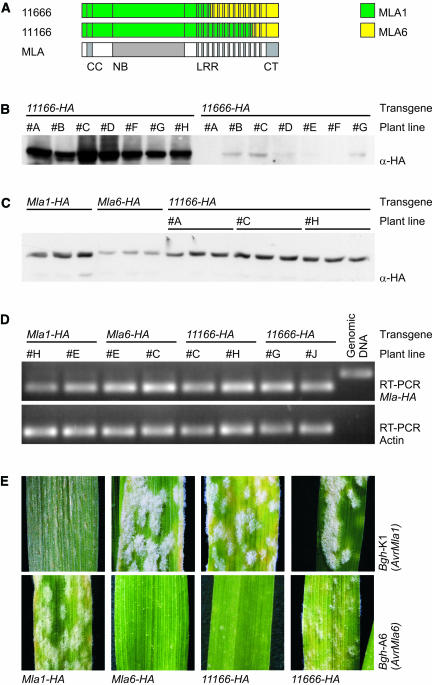
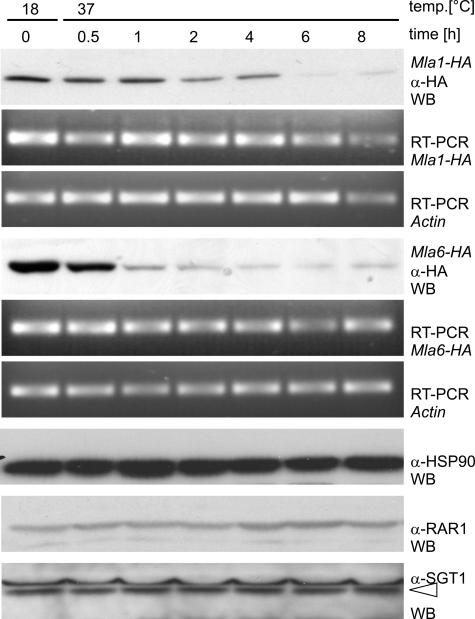
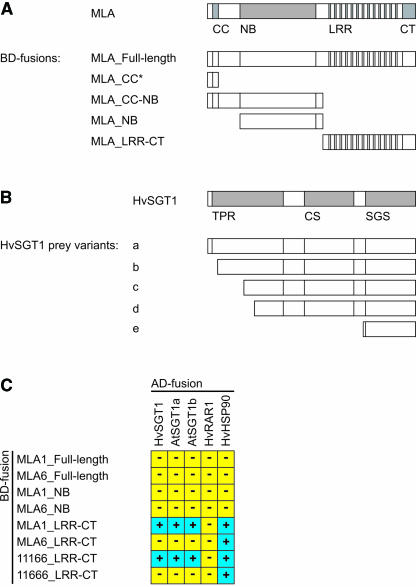
The polymorphic barley (Hordeum vulgare) Mla locus harbors allelic race-specific resistance (R) genes to the powdery mildew fungus Blumeria graminis f sp hordei. The highly sequence-related MLA proteins contain an N-terminal coiled-coil structure, a central nucleotide binding (NB) site, a Leu-rich repeat (LRR) region, and a C-terminal non-LRR region. Using transgenic barley lines expressing epitope-tagged MLA1 and MLA6 derivatives driven by native regulatory sequences, we show a reversible and salt concentration-dependent distribution of the intracellular MLA proteins in soluble and membrane-associated pools. A posttranscriptional process directs fourfold greater accumulation of MLA1 over MLA6. Unexpectedly, in rar1 mutant plants that are compromised for MLA6 but not MLA1 resistance, the steady state level of both MLA isoforms is reduced. Furthermore, differential steady state levels of MLA1/MLA6 hybrid proteins correlate with their requirement for RAR1; the RAR1-independent hybrid protein accumulates to higher levels and the RAR1-dependent one to lower levels. Interestingly, yeast two-hybrid studies reveal that the LRR domains of RAR1-independent but not RAR1-dependent MLA isoforms interact with SGT1, a RAR1 interacting protein required for the function of many NB-LRR type R proteins. Our findings implicate the existence of a conserved mechanism to reach minimal NB-LRR R protein thresholds that are needed to trigger effective resistance responses.
Figures
References
-
- Aust, H.J. (1974). The influence of temperature on infection by powdery mildew Erysiphe graminis f. sp. hordei. Z. Pflanzenkr. Pflanzenschutz 81, 597–601.
-
- Austin, M.J., Muskett, P., Kahn, K., Feys, B.J., Jones, J.D.G., and Parker, J.E. (2002). Regulatory role of SGT1 in early R gene-mediated plant defenses. Science 295, 2077–2080. - PubMed
-
- Axtell, M.J., and Staskawicz, B.J. (2003). Initiation of RPS2-specified disease resistance in Arabidopsis is coupled to the AvrRpt2-directed elimination of RIN4. Cell 112, 369–377. - PubMed
-
- Azevedo, C., Sadanandom, A., Kitagawa, K., Freialdenhoven, A., Shirasu, K., and Schulze-Lefert, P. (2002). The RAR1 interactor SGT1, an essential component of R gene-triggered disease resistance. Science 295, 2073–2076. - PubMed
-
- Bendahmane, A., Farnham, G., Moffett, P., and Baulcombe, D.C. (2002). Constitutive gain-of-function mutants in a nucleotide binding site-leucine rich repeat protein encoded at the Rx locus of potato. Plant J. 32, 195–204. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
