

The role of [Delta]1-pyrroline-5-carboxylate dehydrogenase in proline degradation
- PMID: 15548746
- PMCID: PMC535882
- DOI: 10.1105/tpc.104.023622
The role of [Delta]1-pyrroline-5-carboxylate dehydrogenase in proline degradation
Abstract
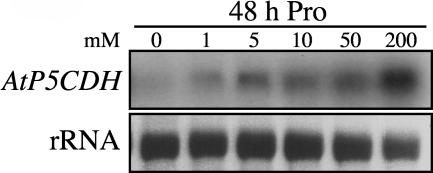
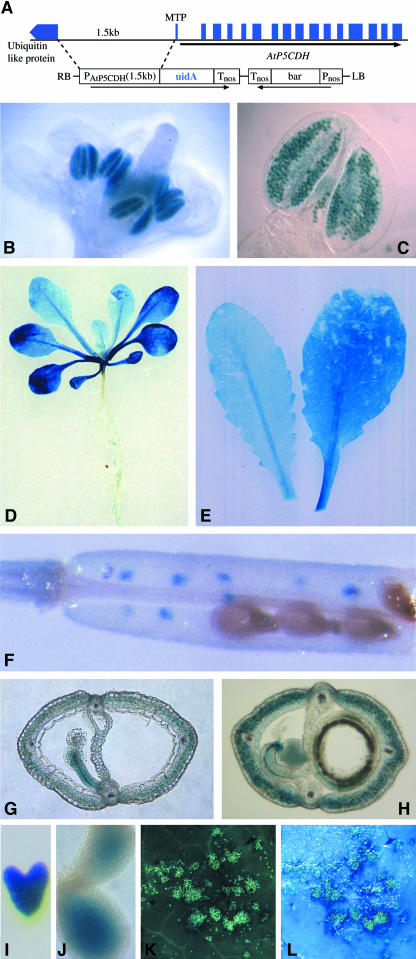
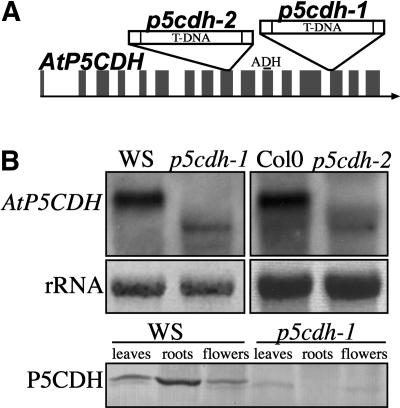
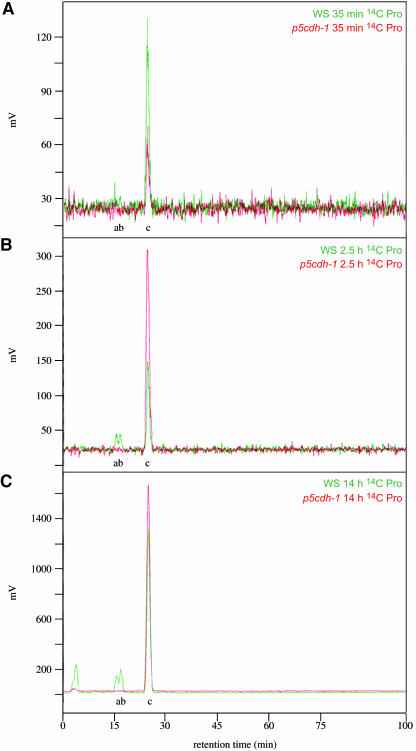
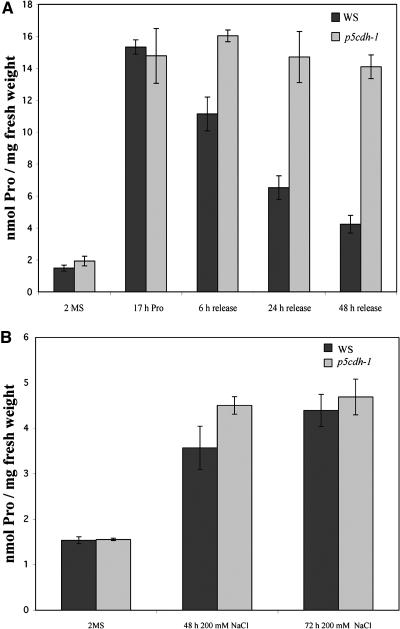
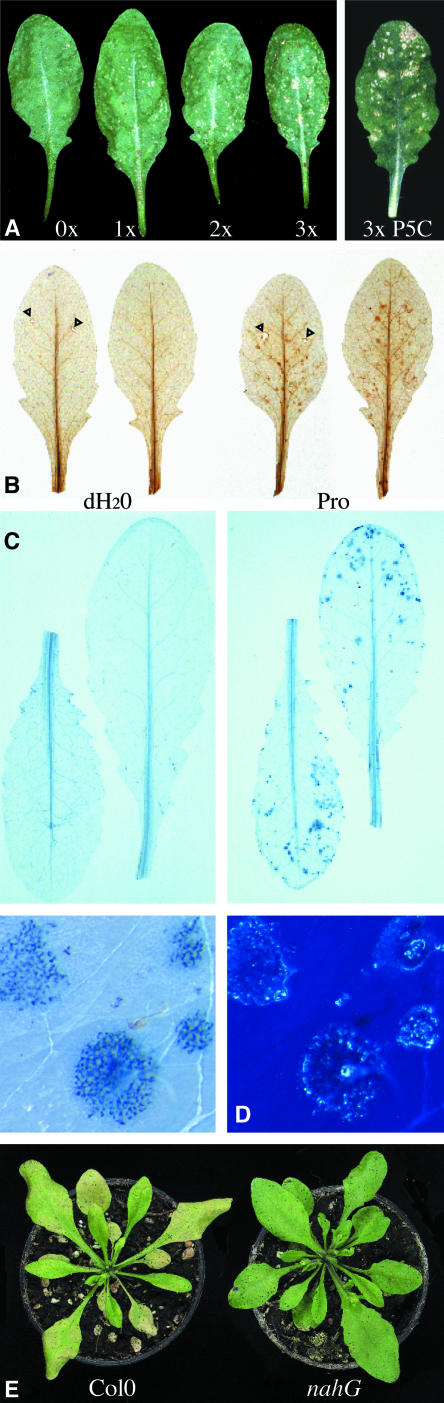
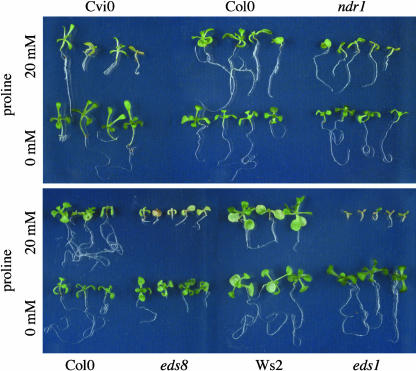
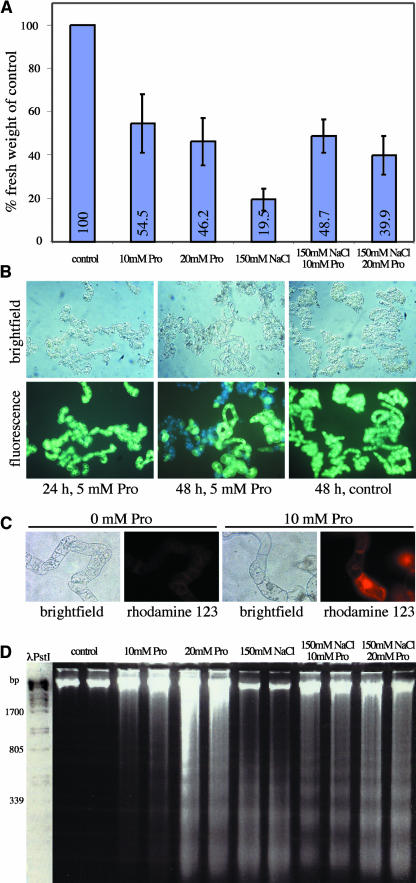
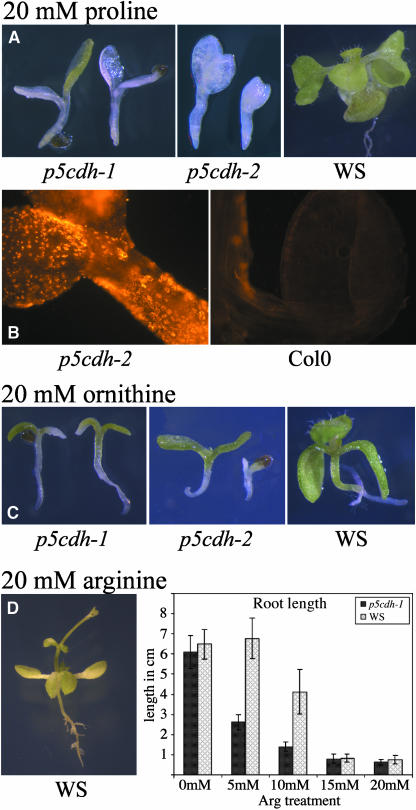
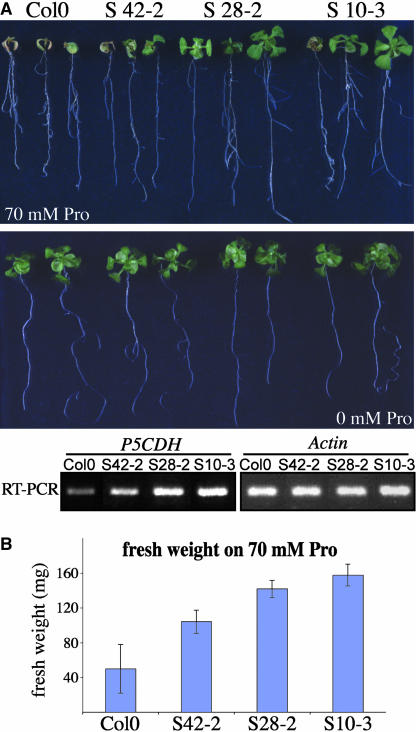
In response to stress, plants accumulate Pro, requiring degradation after release from adverse conditions. Delta1-Pyrroline-5-carboxylate dehydrogenase (P5CDH), the second enzyme for Pro degradation, is encoded by a single gene expressed ubiquitously. To study the physiological function of P5CDH, T-DNA insertion mutants in AtP5CDH were isolated and characterized. Although Pro degradation was undetectable in p5cdh mutants, neither increased Pro levels nor an altered growth phenotype were observed under normal conditions. Thus AtP5CDH is essential for Pro degradation but not required for vegetative plant growth. External Pro application caused programmed cell death, with callose deposition, reactive oxygen species production, and DNA laddering, involving a salicylic acid signal transduction pathway. p5cdh mutants were hypersensitive toward Pro and other molecules producing P5C, such as Arg and Orn. Pro levels were the same in the wild type and mutants, but P5C was detectable only in p5cdh mutants, indicating that P5C accumulation may be the cause for Pro hypersensitivity. Accordingly, overexpression of AtP5CDH resulted in decreased sensitivity to externally supplied Pro. Thus, Pro and P5C/Glu semialdehyde may serve as a link between stress responses and cell death.
Figures
References
-
- Alonso, J.M., et al. (2003). Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301, 653–657. - PubMed
-
- Bates, L.S., Waldren, R.P., and Teare, I.D. (1973). Rapid determination of free proline for water-stress studies. Plant Soil 39, 205–207.
-
- Bonner, C.A., Williams, D.S., Aldrich, H.C., and Jensen, R.A. (1996). Antagonism by L-glutamine of toxicity and growth inhibition caused by other amino acids in suspension cultures of Nicotiana silvestris. Plant Sci. 113, 43–58.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
