

Activation of the complement system generates antibacterial peptides
- PMID: 15550543
- PMCID: PMC534732
- DOI: 10.1073/pnas.0406678101
Activation of the complement system generates antibacterial peptides
Abstract
The complement system represents an evolutionary old and significant part of the innate immune system involved in protection against invading microorganisms. Here, we show that the anaphylatoxin C3a and its inactivated derivative C3a-desArg are antibacterial, demonstrating a previously unknown direct antimicrobial effect of complement activation. The C3a peptide, as well as functional epitopes in the sequence, efficiently killed the Gram-negative bacteria Escherichia coli, Pseudomonas aeruginosa, and the Gram-positive Enterococcus faecalis. In mice, a C3a-derived peptide suppressed infection by Gram-positive Streptococcus pyogenes bacteria. Fluorescence and electron microscopy demonstrated that C3a binds to and induces breaks in bacterial membranes. C3a was also found to induce membrane leakage of liposomes. These findings provide an interesting link between the complement system and antimicrobial peptides, which are two important branches of innate immunity.
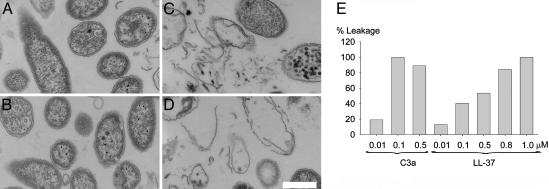
Figures
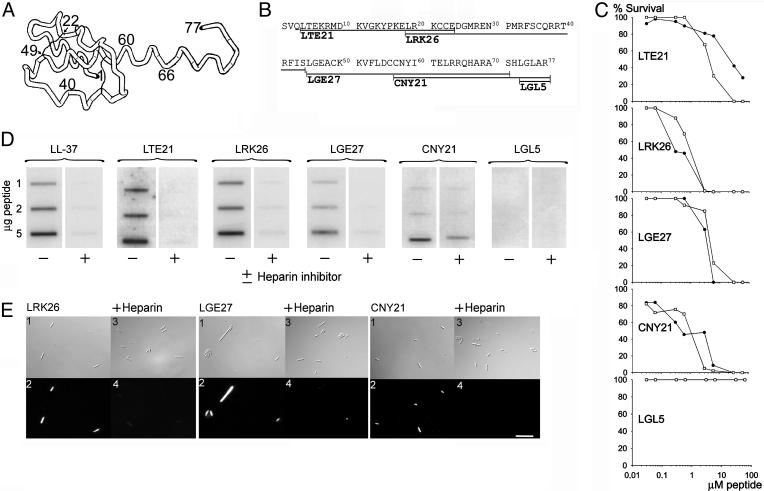
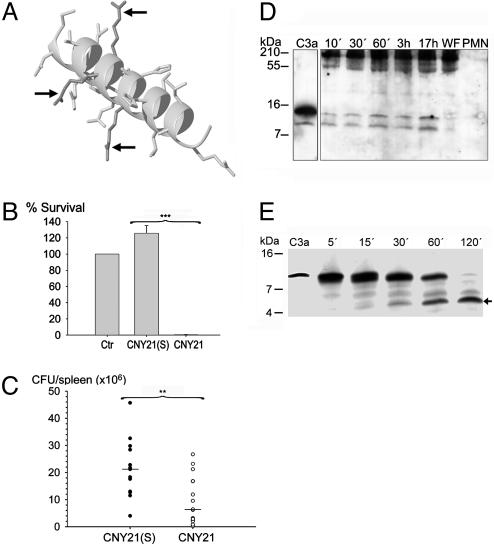
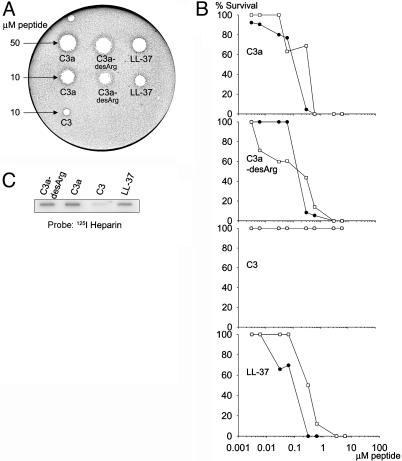
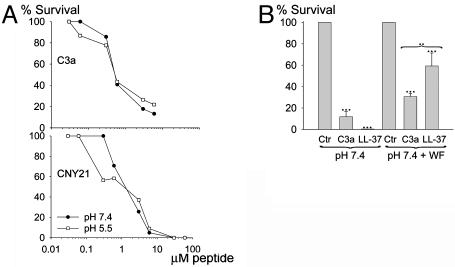
References
-
- Fearon, D. T. & Locksley, R. M. (1996) Science 272, 50-53. - PubMed
-
- Steiner, H., Hultmark, D., Engström, A., Bennich, H. & Boman, H. G. (1981) Nature 292, 246-248. - PubMed
-
- Schröder, J. M. & Harder, J. (1999) Int. J. Biochem. Cell Biol. 31, 645-651. - PubMed
-
- Boman, H. G. (2000) Immunol. Rev. 173, 5-16. - PubMed
-
- Zasloff, M. (2002) Nature 415, 389-395. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
