

A potential role for ICP, a Leishmanial inhibitor of cysteine peptidases, in the interaction between host and parasite
- PMID: 15554964
- PMCID: PMC2244714
- DOI: 10.1111/j.1365-2958.2004.04355.x
A potential role for ICP, a Leishmanial inhibitor of cysteine peptidases, in the interaction between host and parasite
Abstract
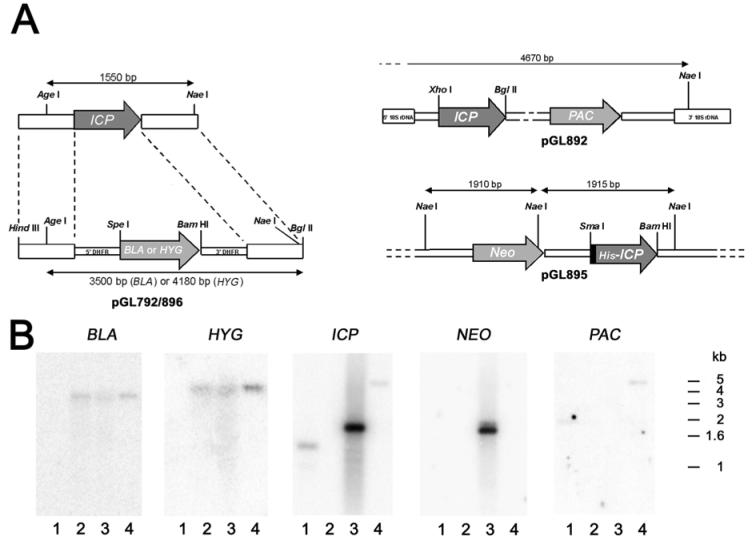
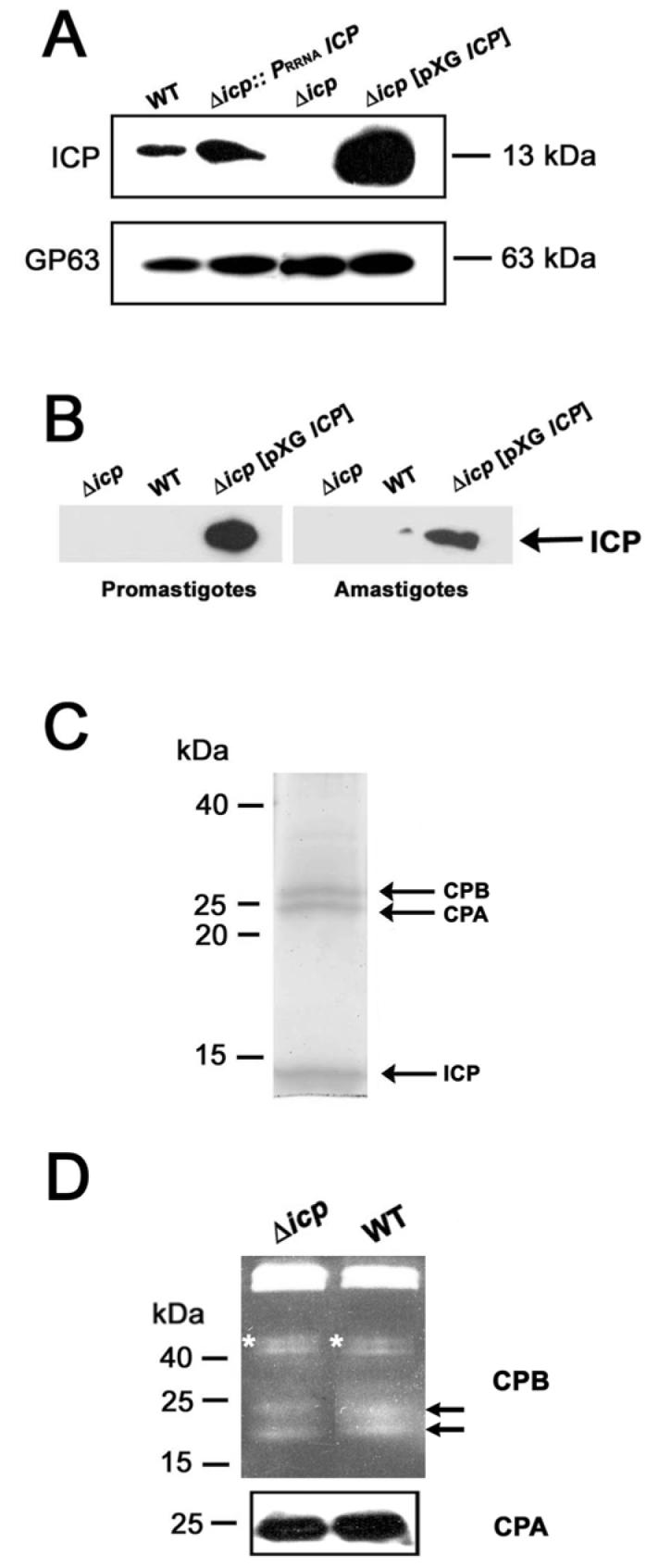
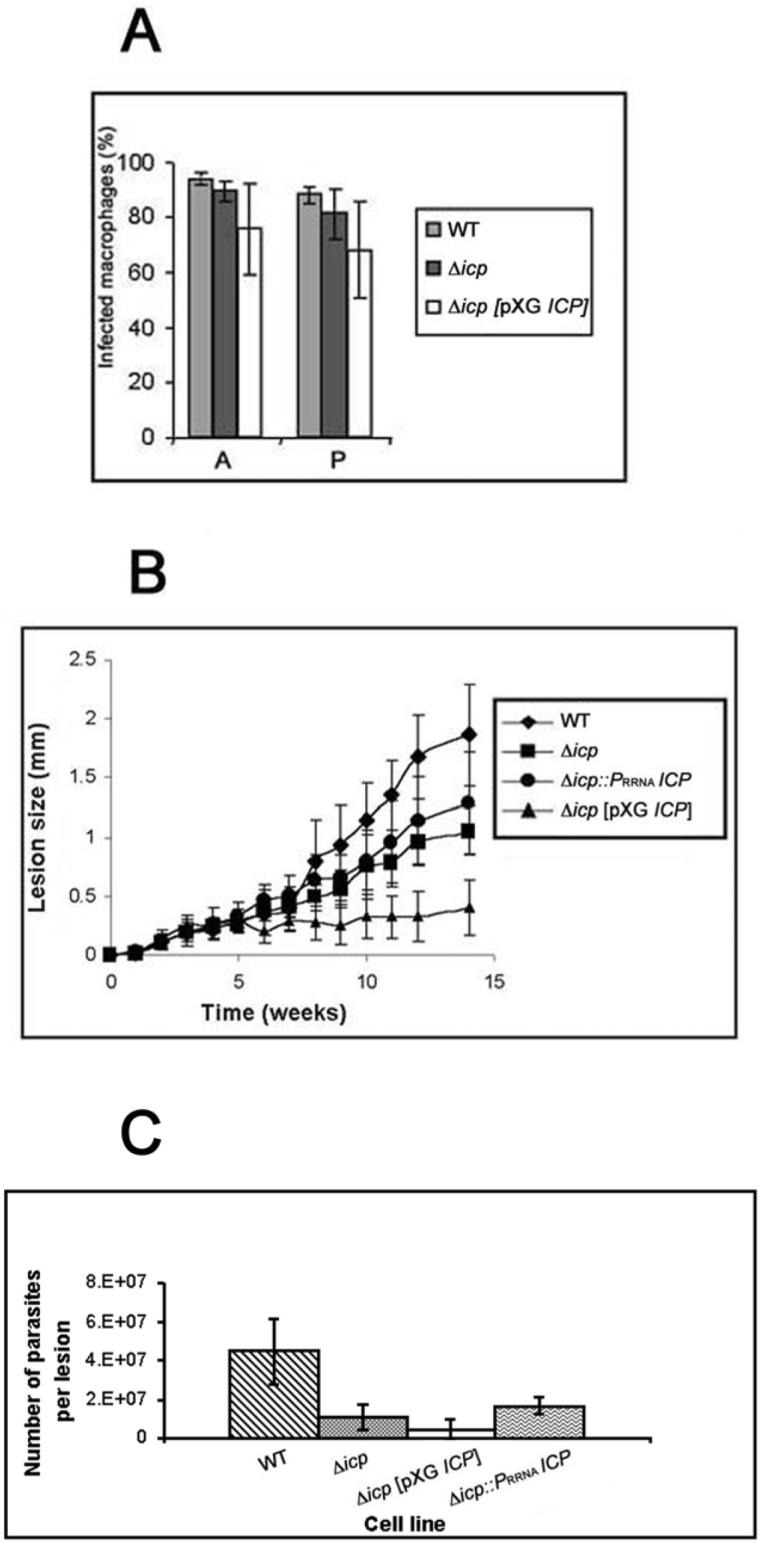
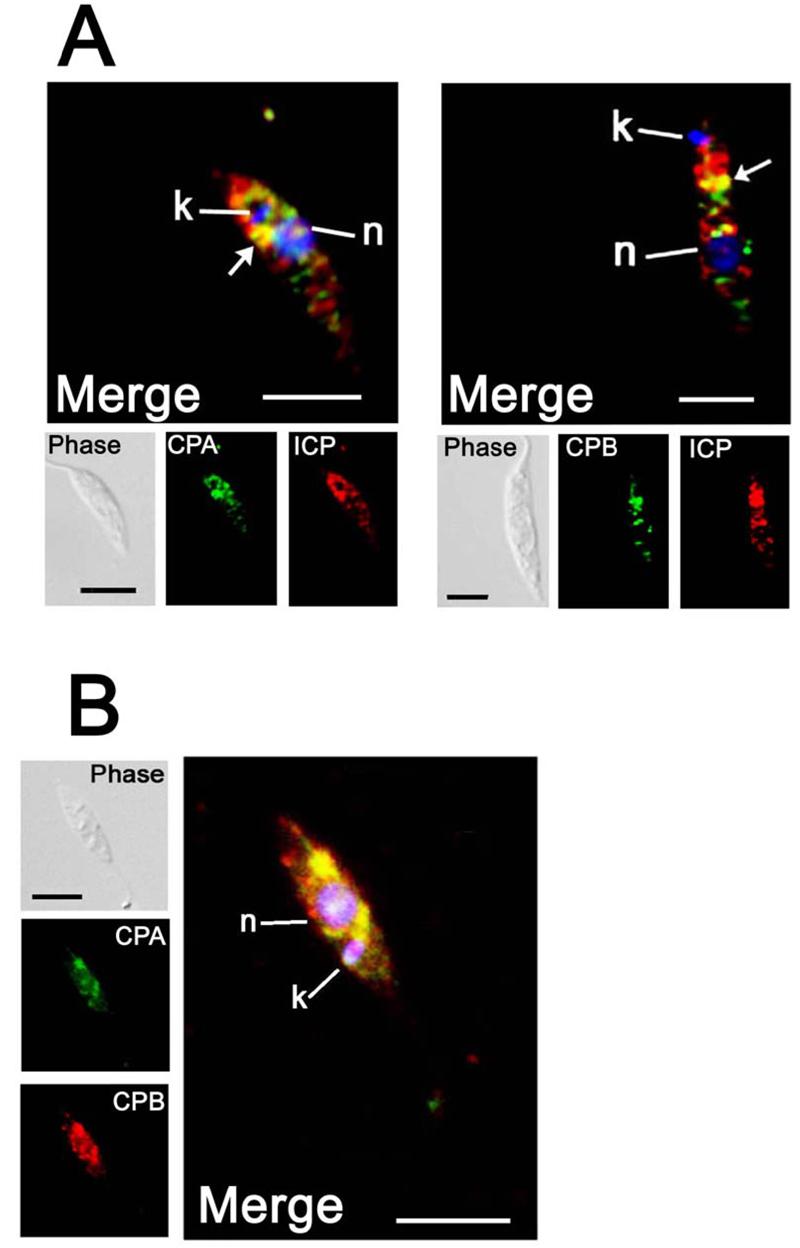
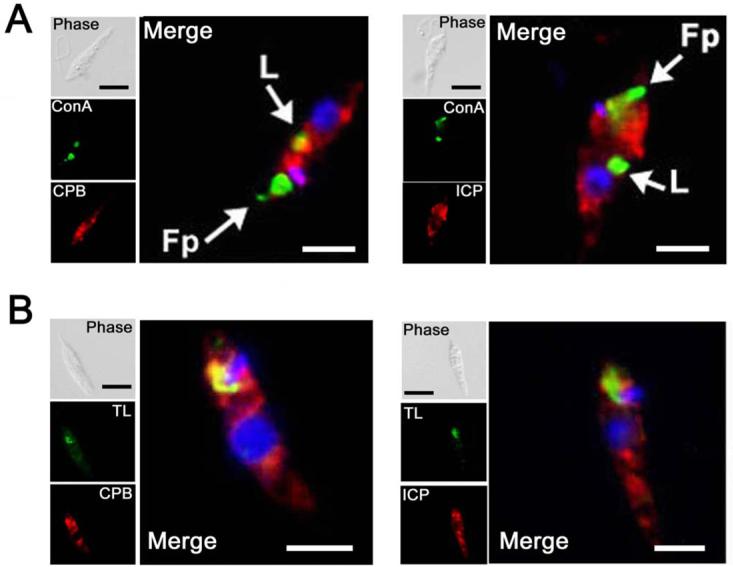
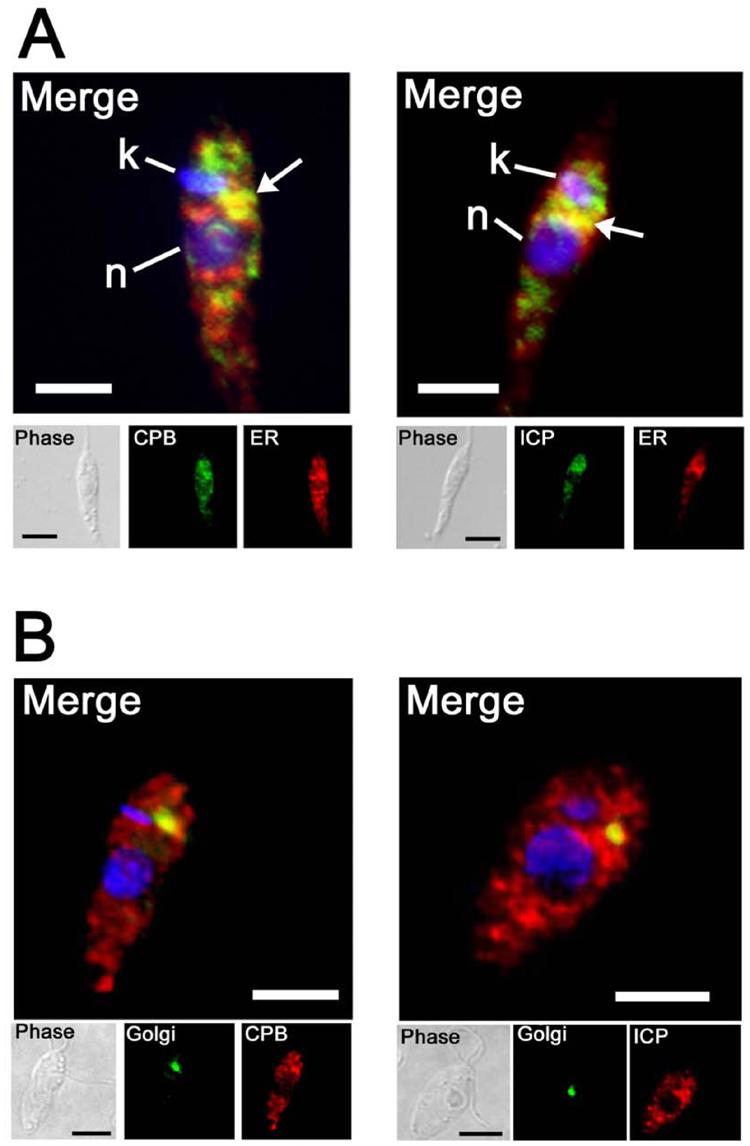
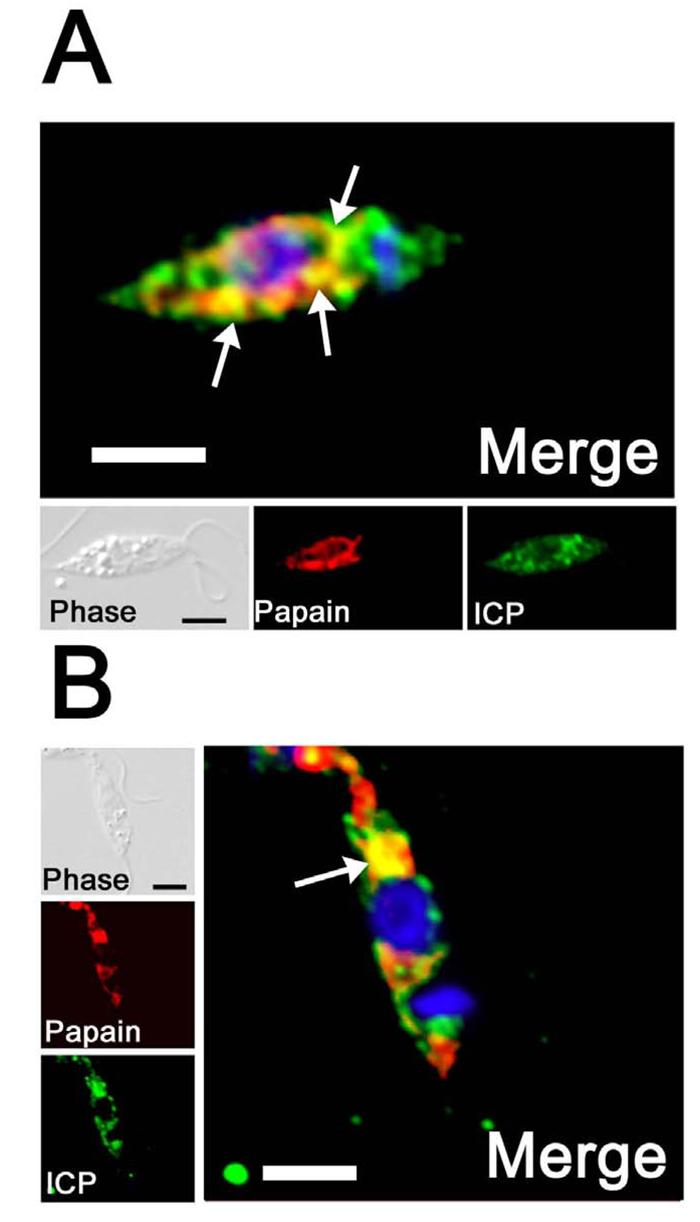
The biological role of a natural inhibitor of cysteine peptidases (designated ICP) of Leishmania has been investigated by genetic manipulation of the parasite. Null mutants grew normally in vitro, were as infective to macrophages in vitro as wild-type parasites, but had reduced infectivity to mice. Mutants re-expressing ICP from a single gene gave partial restoration of virulence in vivo, whereas mutants overexpressing ICP secreted the inhibitor and showed markedly reduced virulence in mice. Promastigotes of the null mutants had similar cysteine peptidase activities as the wild-type parasites, suggesting that ICP is not required for the expression or processing of the enzymes. The only proteins found to bind to ICP in promastigote cell lysates were fully processed forms of CPA and CPB, showing that ICP does not bind in abundance either to zymogens of the cysteine peptidases or other leishmanial proteins. However, only a small proportion of ICP colocalized with CPA and CPB in the promastigote (in the endoplasmic reticulum and Golgi) and the majority of ICP resided in vesicles that are apparently distinct from endosomes and the multivesicular tubule (MVT)-lysosome. These data suggest that ICP has a role other than modulation of the activity of the parasite's own cysteine peptidases and their normal trafficking to the MVT-lysosome via the flagellar pocket. The finding that ICP partially colocalized with an endocytosed cysteine peptidase leads us to postulate that ICP has a role in protection of the parasite against the hydrolytic environment of the sandfly gut and/or the parasitophorous vacuole of host macrophages.
Figures
Similar articles
-
Overexpression of the natural inhibitor of cysteine peptidases in Leishmania mexicana leads to reduced virulence and a Th1 response.Infect Immun. 2009 Jul;77(7):2971-8. doi: 10.1128/IAI.00558-08. Epub 2009 May 11. Infect Immun. 2009. PMID: 19433541 Free PMC article.
-
Leishmania mexicana cysteine proteinase-deficient mutants have attenuated virulence for mice and potentiate a Th1 response.J Immunol. 1998 Dec 15;161(12):6794-801. J Immunol. 1998. PMID: 9862710
-
Inhibitor of cysteine peptidase does not influence the development of Leishmania mexicana in Lutzomyia longipalpis.J Med Entomol. 2009 May;46(3):605-9. doi: 10.1603/033.046.0327. J Med Entomol. 2009. PMID: 19496433
-
Cysteine peptidases as virulence factors of Leishmania.Curr Opin Microbiol. 2004 Aug;7(4):375-81. doi: 10.1016/j.mib.2004.06.010. Curr Opin Microbiol. 2004. PMID: 15358255 Review.
-
Cysteine Proteases Inhibitors with Immunoglobulin-Like Fold in Protozoan Parasites and their Role in Pathogenesis.Curr Protein Pept Sci. 2017;18(10):1035-1042. doi: 10.2174/1389203717666160813163837. Curr Protein Pept Sci. 2017. PMID: 27526930 Review.
Cited by
-
Endocytosis and Sphingolipid Scavenging in Leishmania mexicana Amastigotes.Biochem Res Int. 2012;2012:691363. doi: 10.1155/2012/691363. Epub 2011 Sep 21. Biochem Res Int. 2012. PMID: 21941657 Free PMC article.
-
Influence of parasite encoded inhibitors of serine peptidases in early infection of macrophages with Leishmania major.Cell Microbiol. 2009 Jan;11(1):106-20. doi: 10.1111/j.1462-5822.2008.01243.x. Epub 2008 Oct 29. Cell Microbiol. 2009. PMID: 19016791 Free PMC article.
-
The structure of Leishmania mexicana ICP provides evidence for convergent evolution of cysteine peptidase inhibitors.J Biol Chem. 2006 Mar 3;281(9):5821-8. doi: 10.1074/jbc.M510868200. Epub 2005 Dec 28. J Biol Chem. 2006. PMID: 16407198 Free PMC article.
-
Protein turnover and differentiation in Leishmania.Int J Parasitol. 2007 Aug;37(10):1063-75. doi: 10.1016/j.ijpara.2007.03.008. Epub 2007 Mar 31. Int J Parasitol. 2007. PMID: 17493624 Free PMC article. Review.
-
Ecotin-like serine peptidase inhibitor ISP1 of Leishmania major plays a role in flagellar pocket dynamics and promastigote differentiation.Cell Microbiol. 2012 Aug;14(8):1271-86. doi: 10.1111/j.1462-5822.2012.01798.x. Epub 2012 May 8. Cell Microbiol. 2012. PMID: 22486816 Free PMC article.
References
-
- Abrahamson M, Alvarez-Fernandez M, Nathanson CM. Cystatins. Biochemical Society Symposium. 2003;70:179–199. - PubMed
-
- Alexander J, Coombs GH, Mottram JC. Leishmania mexicana cysteine proteinase-deficient mutants have attenuated virulence for mice and potentiate a T Helper 1 (TH1) response. J Immunol. 1998;161:6794–6801. - PubMed
-
- Alexander J, Satoskar AR, Russell DG. Leishmania species: models of intracellular parasitism. J Cell Sci. 1999;112:2993–3002. - PubMed
-
- Bart G, Coombs GH, Mottram JC. Isolation of lmcpc, a gene encoding a Leishmania mexicana cathepsin B-like cysteine proteinase. Mol Biochem Parasitol. 1995;73:271–274. - PubMed
-
- Bates PA, Robertson CD, Tetley L, Coombs GH. Axenic cultivation and characterization of Leishmania mexicana amastigote-like forms. Parasitology. 1992;105:193–202. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
