

Tumor cell traffic through the extracellular matrix is controlled by the membrane-anchored collagenase MT1-MMP
- PMID: 15557125
- PMCID: PMC2172570
- DOI: 10.1083/jcb.200408028
Tumor cell traffic through the extracellular matrix is controlled by the membrane-anchored collagenase MT1-MMP
Abstract
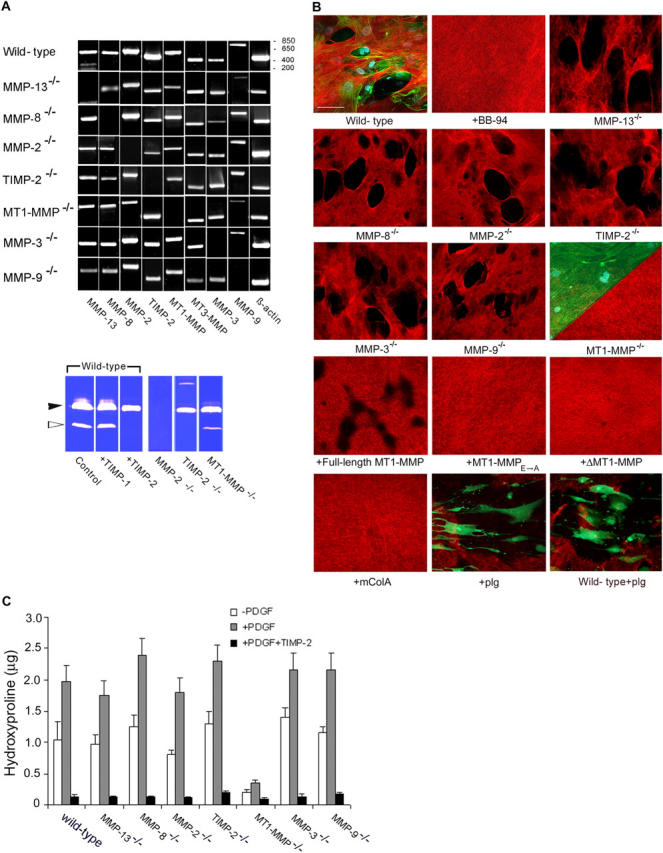
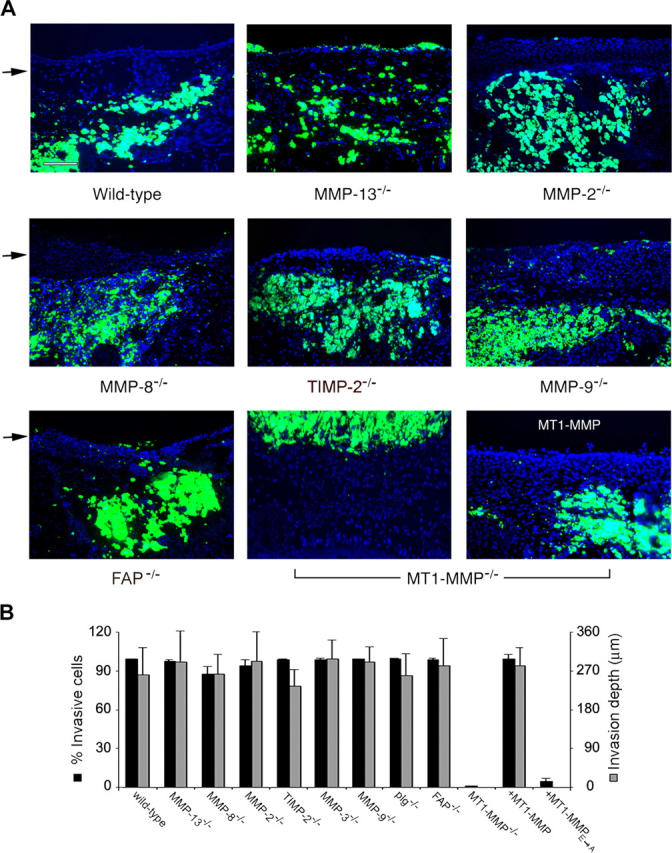
As cancer cells traverse collagen-rich extracellular matrix (ECM) barriers and intravasate, they adopt a fibroblast-like phenotype and engage undefined proteolytic cascades that mediate invasive activity. Herein, we find that fibroblasts and cancer cells express an indistinguishable pericellular collagenolytic activity that allows them to traverse the ECM. Using fibroblasts isolated from gene-targeted mice, a matrix metalloproteinase (MMP)-dependent activity is identified that drives invasion independently of plasminogen, the gelatinase A/TIMP-2 axis, gelatinase B, collagenase-3, collagenase-2, or stromelysin-1. In contrast, deleting or suppressing expression of the membrane-tethered MMP, MT1-MMP, in fibroblasts or tumor cells results in a loss of collagenolytic and invasive activity in vitro or in vivo. Thus, MT1-MMP serves as the major cell-associated proteinase necessary to confer normal or neoplastic cells with invasive activity.
Figures




References
-
- Armstrong, P.B., J.P. Quigley, and E. Sidebottom. 1982. Transepithelial invasion and intramesenchymal infiltration of the chick embryo chorioallantois by tumor cell lines. Cancer Res. 42:1826–1837. - PubMed
-
- Balbin, M., A. Fueyo, V. Knauper, J.M. Lopez, J. Alvarez, L.M. Sanchez, V. Quesada, J. Bordallo, G. Murphy, and V. Lopez-Otin. 2001. Identification and enzymatic characterization of two diverging murine counterparts of human interstitial collagenase (MMP-1) expressed at sites of embryo implantation. J. Biol. Chem. 276:10253–10262. - PubMed
-
- Balbin, M., A. Fueyo, A.M. Tester, A.M. Pendas, A.S. Pitiot, A. Astudillo, C.M. Overall, S.D. Shapiro, and C. Lopez-Otin. 2003. Loss of collagen-2 confers increased skin tumor susceptibility to male mice. Nat. Genet. 35:252–257. - PubMed
-
- Benbow, U., M.P. Schoenermark, T.I. Mitchell, J.L. Rutter, K. Shimokawa, H. Nagase, and C.E. Brinckerhoff. 1999. A novel host/tumor cell interaction activates matrix metalloproteinase 1 and mediates invasion through type I collagen. J. Biol. Chem. 274:25371–25378. - PubMed
-
- Birkedal-Hansen, H. 1987. Catabolism and turnover of collagens: collagenases. Methods Enzymol. 144:140–171. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
