

Structural basis for the restoration of TCR recognition of an MHC allelic variant by peptide secondary anchor substitution
- PMID: 15557346
- PMCID: PMC2211956
- DOI: 10.1084/jem.20040217
Structural basis for the restoration of TCR recognition of an MHC allelic variant by peptide secondary anchor substitution
Abstract
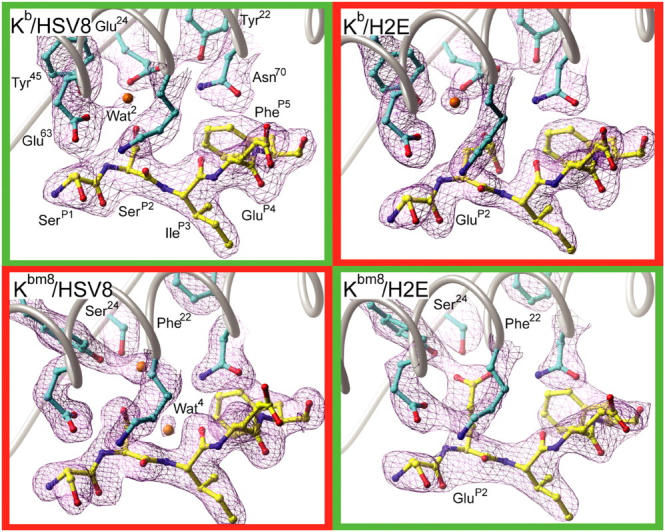
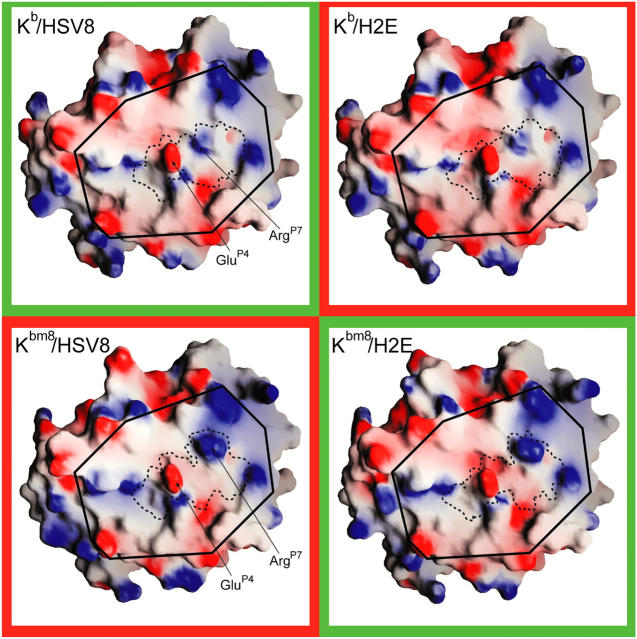
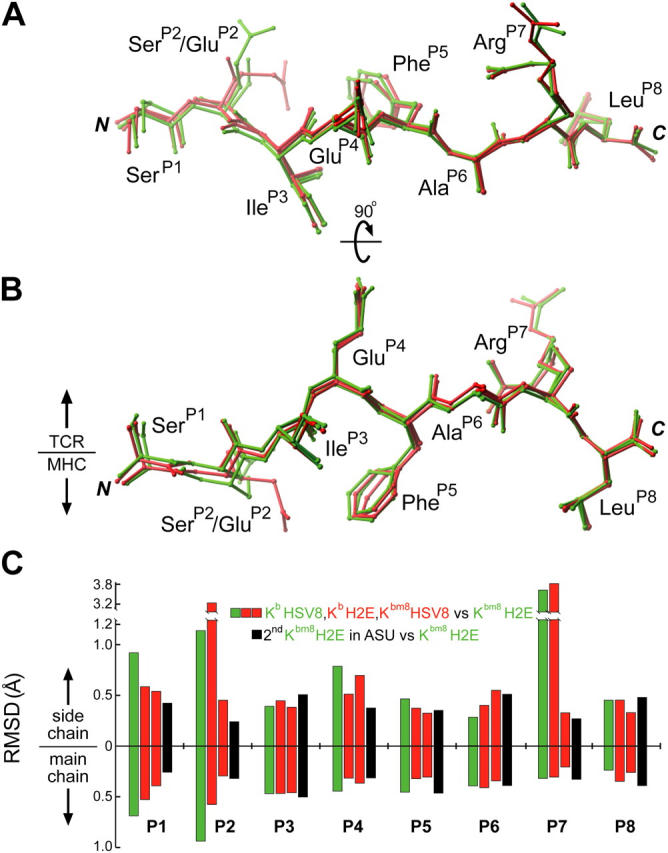
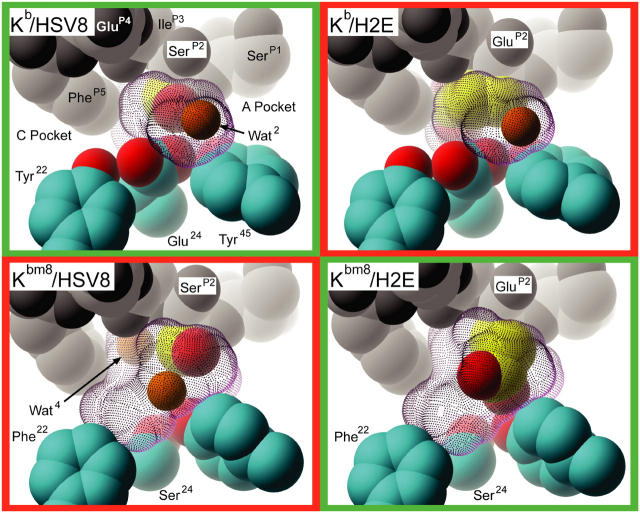
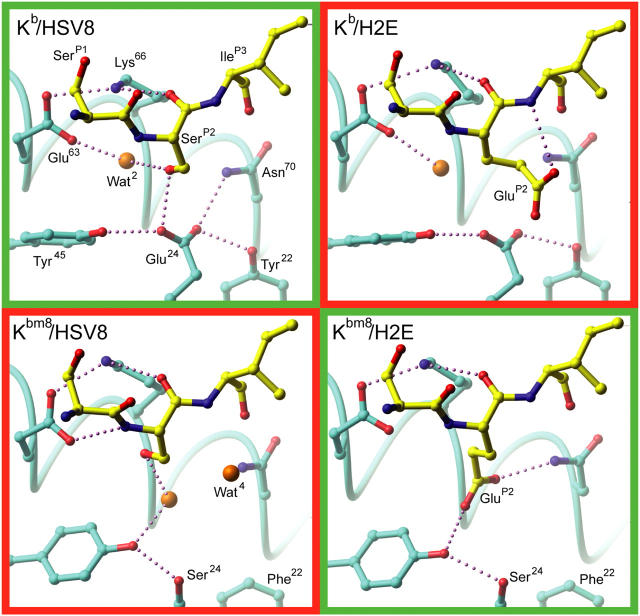
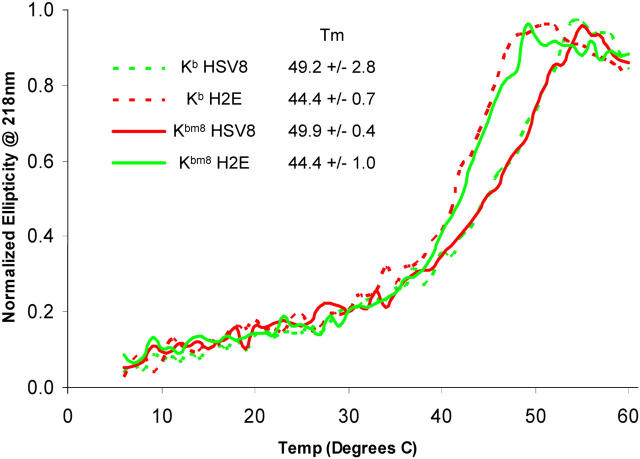
Major histocompatibility complex (MHC) class I variants H-2K(b) and H-2K(bm8) differ primarily in the B pocket of the peptide-binding groove, which serves to sequester the P2 secondary anchor residue. This polymorphism determines resistance to lethal herpes simplex virus (HSV-1) infection by modulating T cell responses to the immunodominant glycoprotein B(498-505) epitope, HSV8. We studied the molecular basis of these effects and confirmed that T cell receptors raised against K(b)-HSV8 cannot recognize H-2K(bm8)-HSV8. However, substitution of Ser(P2) to Glu(P2) (peptide H2E) reversed T cell receptor (TCR) recognition; H-2K(bm8)-H2E was recognized whereas H-2K(b)-H2E was not. Insight into the structural basis of this discrimination was obtained by determining the crystal structures of all four MHC class I molecules in complex with bound peptide (pMHCs). Surprisingly, we find no concerted pMHC surface differences that can explain the differential TCR recognition. However, a correlation is apparent between the recognition data and the underlying peptide-binding groove chemistry of the B pocket, revealing that secondary anchor residues can profoundly affect TCR engagement through mechanisms distinct from the alteration of the resting state conformation of the pMHC surface.
Figures
References
-
- Matsumura, M., D.H. Fremont, P.A. Peterson, and I.A. Wilson. 1992. Emerging principles for the recognition of peptide antigens by MHC class I molecules. Science. 257:927–934. - PubMed
-
- Madden, D.R. 1995. The three-dimensional structure of peptide-MHC complexes. Annu. Rev. Immunol. 13:587–622. - PubMed
-
- Bjorkman, P.J., M.A. Saper, B. Samraoui, W.S. Bennett, J.L. Strominger, and D.C. Wiley. 1987. The foreign antigen binding site and T cell recognition regions of class I histocompatibility antigens. Nature. 329:512–518. - PubMed
-
- Rammensee, H.G., K. Falk, and O. Rotzschke. 1993. Peptides naturally presented by MHC class I molecules. Annu. Rev. Immunol. 11:213–244. - PubMed
-
- van der Merwe, P.A., and S.J. Davis. 2003. Molecular interactions mediating T cell antigen recognition. Annu. Rev. Immunol. 21:659–684. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
