

Extreme heterogeneity in the molecular events leading to the establishment of chiasmata during meiosis i in human oocytes
- PMID: 15558497
- PMCID: PMC1196414
- DOI: 10.1086/427268
Extreme heterogeneity in the molecular events leading to the establishment of chiasmata during meiosis i in human oocytes
Abstract
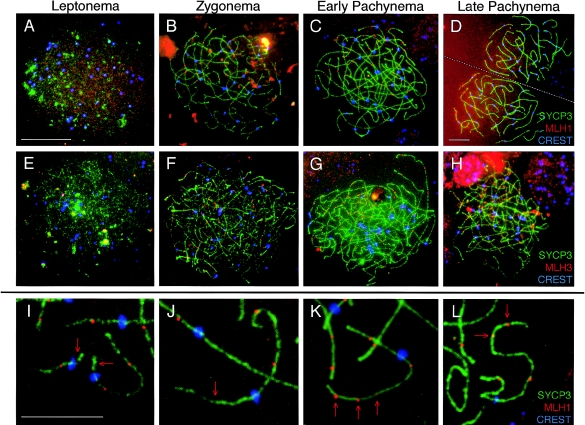
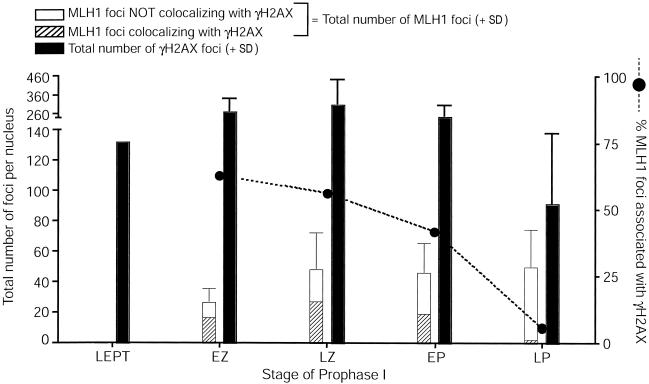
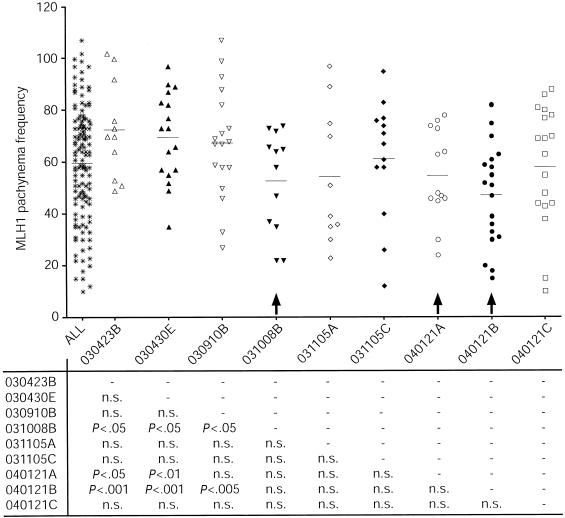
In humans, ~50% of conceptuses are chromosomally aneuploid as a consequence of errors in meiosis, and most of these aneuploid conceptuses result in spontaneous miscarriage. Of these aneuploidy events, 70% originate during maternal meiosis, with the majority proposed to arise as a direct result of defective crossing over during meiotic recombination in prophase I. By contrast, <1%-2% of mouse germ cells exhibit prophase I-related nondisjunction events. This disparity among mammalian species is surprising, given the conservation of genes and events that regulate meiotic progression. To understand the mechanisms that might be responsible for the high error rates seen in human females, we sought to further elucidate the regulation of meiotic prophase I at the molecular cytogenetic level. Given that these events occur during embryonic development in females, samples were obtained during a defined period of gestation (17-24 weeks). Here, we demonstrate that human oocytes enter meiotic prophase I and progress through early recombination events in a similar temporal framework to mice. However, at pachynema, when chromosomes are fully paired, we find significant heterogeneity in the localization of the MutL homologs, MLH1 and MLH3, among human oocyte populations. MLH1 and MLH3 have been shown to mark late-meiotic nodules that correlate well with--and are thought to give rise to--the sites of reciprocal recombination between homologous chromosomes, which suggests a possible 10-fold variation in the processing of nascent recombination events. If such variability persists through development and into adulthood, these data would suggest that as many as 30% of human oocytes are predisposed to aneuploidy as a result of prophase I defects in MutL homolog-related events.
Figures
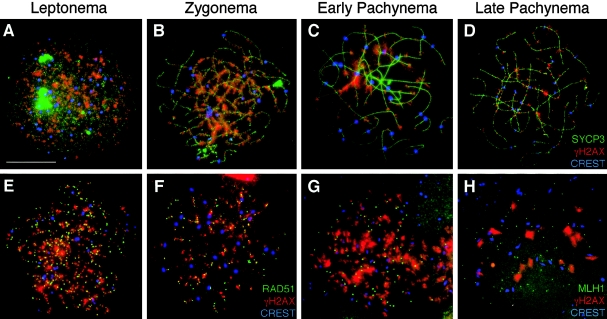
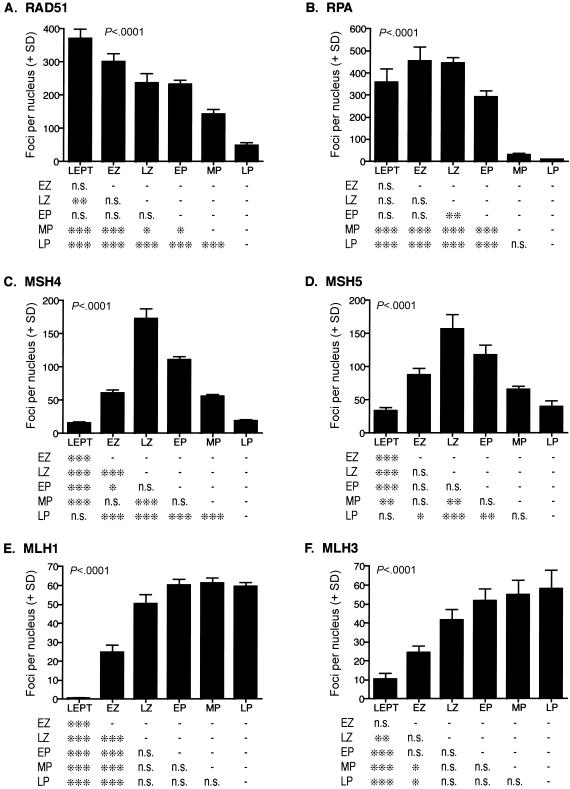
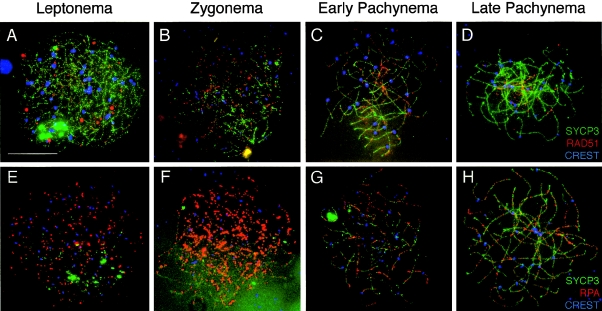
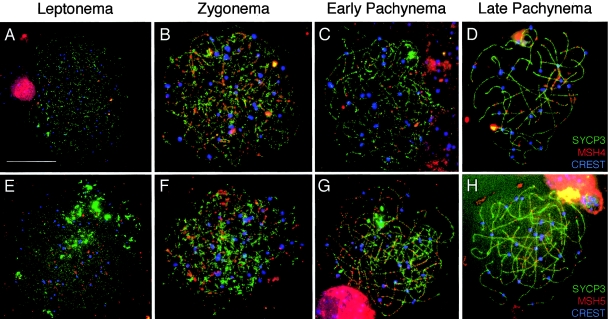
References
Electronic-Database Information
-
- Online Mendelian Inheritance in Man (OMIM), http://www.ncbi.nlm.nih.gov/Omim/ (for Sycp3, RAD51, RPA, MSH4, MSH5, MLH1, MLH3, H2AX, and SPO11 clone)
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
