

Degradation of human antimicrobial peptide LL-37 by Staphylococcus aureus-derived proteinases
- PMID: 15561843
- PMCID: PMC529204
- DOI: 10.1128/AAC.48.12.4673-4679.2004
Degradation of human antimicrobial peptide LL-37 by Staphylococcus aureus-derived proteinases
Abstract
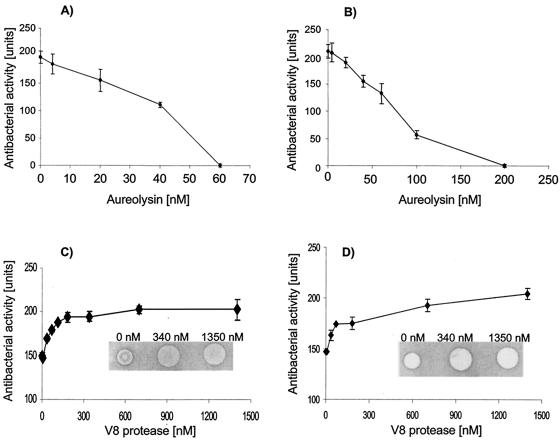
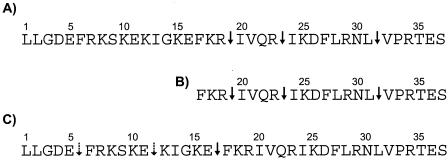
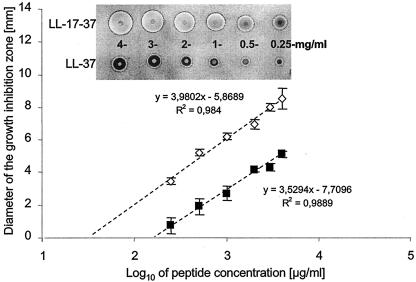
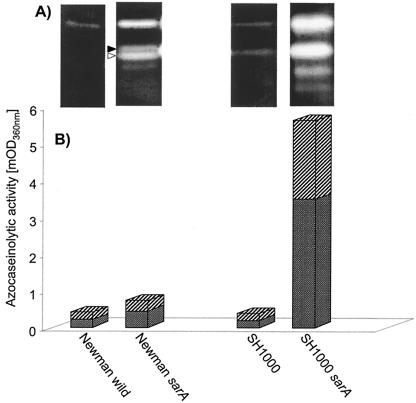
Cathelicidin LL-37 is one of the few human bactericidal peptides with potent antistaphylococcal activity. In this study we examined the susceptibility of LL-37 to proteolytic degradation by two major proteinases produced by Staphylococcus aureus, a metalloproteinase (aureolysin) and a glutamylendopeptidase (V8 protease). We found that aureolysin cleaved and inactivated LL-37 in a time- and concentration-dependent manner. Analysis of the generated fragments by mass spectroscopy revealed that the initial cleavage of LL-37 by aureolysin occurred between the Arg19-Ile20, Arg23-Ile24, and Leu31-Val32 peptide bonds, instantly annihilating the antibacterial activity of LL-37. In contrast, the V8 proteinase hydrolyzed efficiently only the Glu16-Phe17 peptide bond, rendering the C-terminal fragment refractory to further degradation. This fragment (termed LL-17-37) displayed antibacterial activity against S. aureus at a molar level similar to that of the full-length LL-37 peptide, indicating that the antibacterial activity of LL-37 resides in the C-terminal region. In keeping with LL-37 degradation by aureolysin, S. aureus strains that produce significant amounts of this metalloprotease were found to be less susceptible to LL-17-37 than strains expressing no aureolysin activity. Taken together, these data suggest that aureolysin production by S. aureus contributes to the resistance of this pathogen to the innate immune system of humans mediated by LL-37.
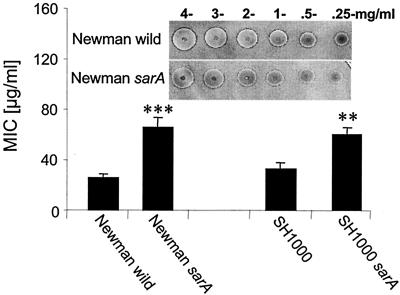
Figures
References
-
- Belaaouaj, A., R. McCarthy, M. Baumann, Z. Gao, T. J. Ley, S. N. Abraham, and S. D. Shapiro. 1998. Mice lacking neutrophil elastase reveal impaired host defense against gram negative bacterial sepsis. Nat. Med. 4:615-618. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
