

The heteromeric organic solute transporter alpha-beta, Ostalpha-Ostbeta, is an ileal basolateral bile acid transporter
- PMID: 15563450
- PMCID: PMC1224727
- DOI: 10.1074/jbc.M412752200
The heteromeric organic solute transporter alpha-beta, Ostalpha-Ostbeta, is an ileal basolateral bile acid transporter
Abstract
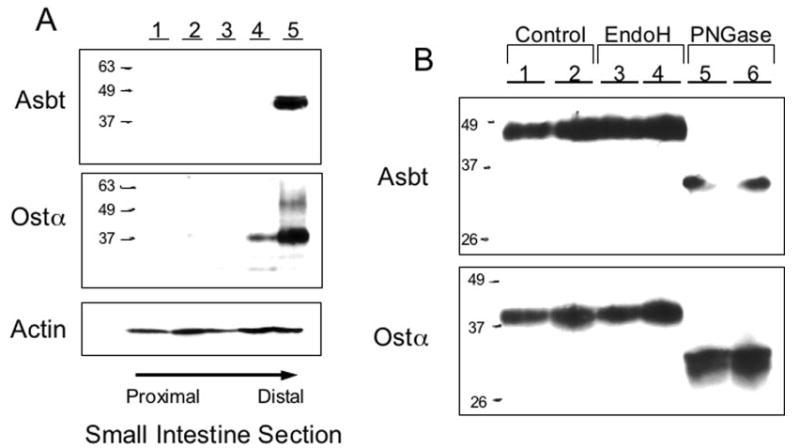
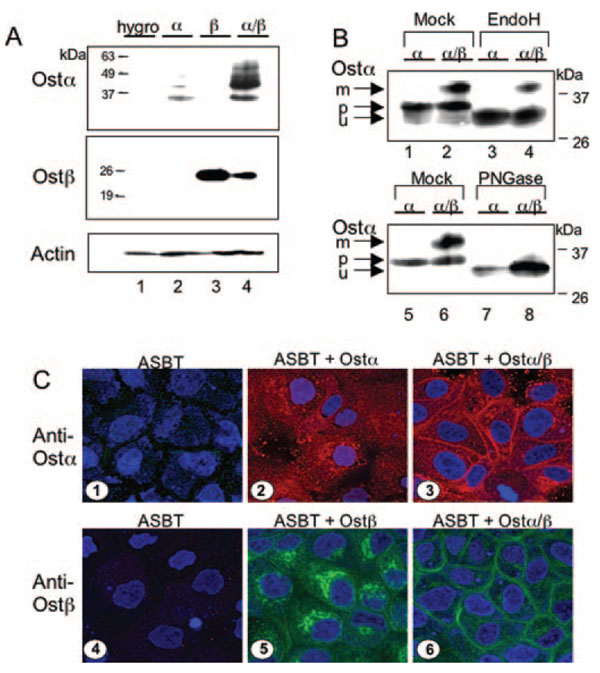
Bile acids are transported across the ileal enterocyte brush border membrane by the well characterized apical sodium-dependent bile acid transporter (Asbt) Slc10a2; however, the carrier(s) responsible for transporting bile acids across the ileocyte basolateral membrane into the portal circulation have not been fully identified. Transcriptional profiling of wild type and Slc10a2 null mice was employed to identify a new candidate basolateral bile acid carrier, the heteromeric organic solute transporter (Ost)alpha-Ostbeta. By Northern blot analysis, Ostalpha and Ostbeta mRNA was detected only in mouse kidney and intestine, mirroring the horizontal gradient of expression of Asbt in the gastrointestinal tract. Analysis of Ostalpha and Ostbeta protein expression by immunohistochemistry localized both subunits to the basolateral surface of the mouse ileal enterocyte. The transport properties of Ostalpha-Ostbeta were analyzed in stably transfected Madin-Darby canine kidney cells. Co-expression of mouse Ostalpha-Ostbeta, but not the individual subunits, stimulated Na(+)-independent bile acid uptake and the apical-to-basolateral transport of taurocholate. In contrast, basolateral-to-apical transport was not affected by Ostalpha-Ostbeta expression. Co-expression of Ostalpha and Ostbeta was required to convert the Ostalpha subunit to a mature glycosylated endoglycosidase H-resistant form, suggesting that co-expression facilitates the trafficking of Ostalpha through the Golgi apparatus. Immunolocalization studies showed that co-expression was necessary for plasma membrane expression of both Ostalpha and Ostbeta. These results demonstrate that the mouse Ostalpha-Ostbeta heteromeric transporter is a basolateral bile acid carrier and may be responsible for bile acid efflux in ileum and other ASBT-expressing tissues.
Figures






References
-
- Shneider, B. L. (2001) J. Pediatr. Gastroenterol. Nutr.32, 407–417 - PubMed
-
- Dawson, P. A., Haywood, J., Craddock, A. L., Wilson, M., Tietjen, M., Kluckman, K., Maeda, N., Parks, J. S. (2003) J. Biol. Chem.278, 33920–33927 - PubMed
-
- Lin, M. C., Weinberg, S. L., Kramer, W., Burckhardt, G., and Wilson, F. A. (1988) J. Membr. Biol.106, 1–11 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
