

Coordinated assembly of human translation initiation complexes by the hepatitis C virus internal ribosome entry site RNA
- PMID: 15563596
- PMCID: PMC534415
- DOI: 10.1073/pnas.0407402101
Coordinated assembly of human translation initiation complexes by the hepatitis C virus internal ribosome entry site RNA
Abstract
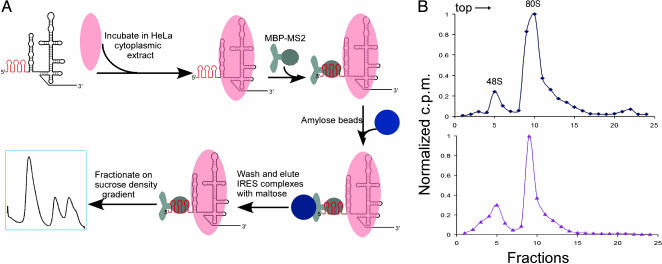
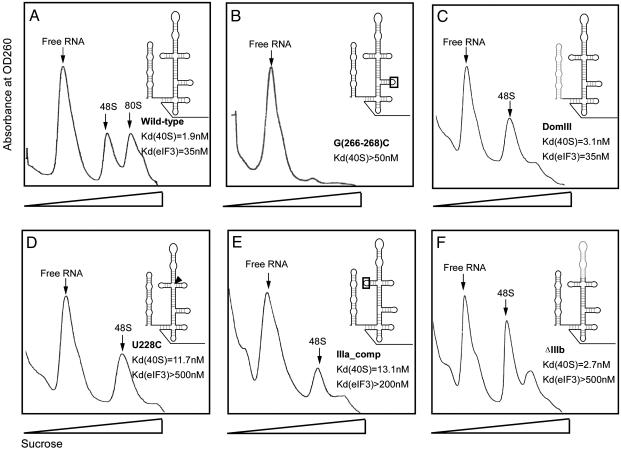
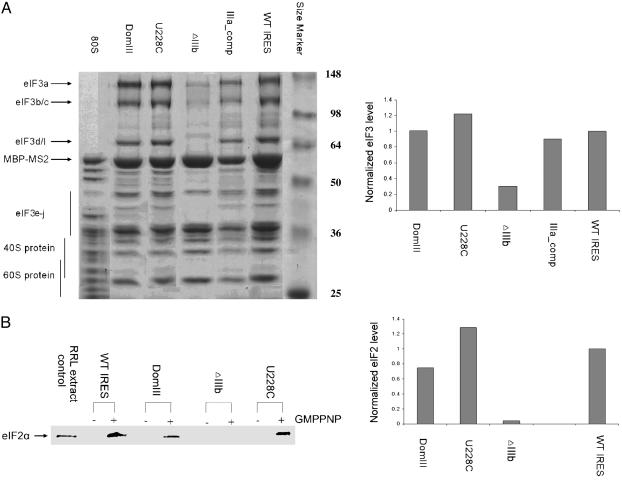
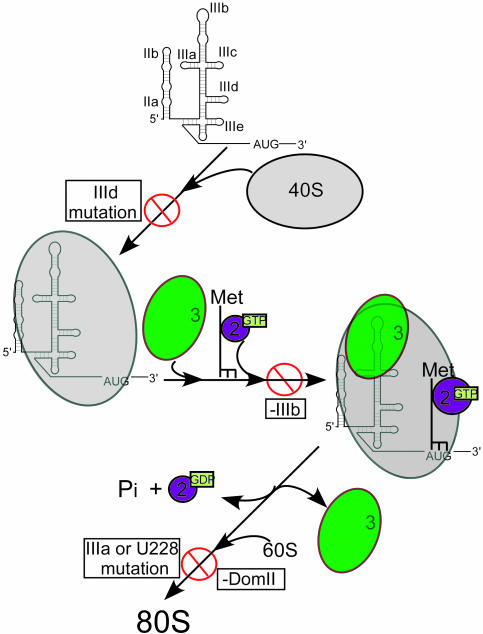
Protein synthesis in all cells begins with recruitment of the small ribosomal subunit to the initiation codon in a messenger RNA. In some eukaryotic viruses, RNA upstream of the coding region forms an internal ribosome entry site (IRES) that directly binds to the 40S ribosomal subunit and enables translation initiation in the absence of many canonical translation initiation factors. The hepatitis C virus (HCV) IRES RNA requires just two initiation factors, eukaryotic initiation factor (eIF) 2 and eIF3, to form preinitiation 48S ribosomal complexes that subsequently assemble into translation-competent ribosomes. Using an RNA-based affinity purification approach, we show here that HCV IRES RNA facilitates eIF2 function through its interactions with eIF3 and the 40S ribosomal subunit. Although the wild-type IRES assembles normally into 48S and 80S ribosomal complexes in human cell extract, mutant IRES RNAs become trapped at the 48S assembly stage. Trapped 48S complexes formed by IRES mutants with reduced eIF3 binding affinity nonetheless contain eIF3, consistent with inherent eIF3-40S subunit affinity. Intriguingly, however, one of these IRES mutants prevents stable association of both eIF3 and eIF2, preventing initiator tRNA deposition and explaining the block in 80S assembly. In contrast, an IRES mutant unable to induce a conformational change in the 40S subunit, as observed previously by single-particle cryoelectron microscopy, blocks 80S formation at a later stage in assembly. These data suggest that the IRES RNA coordinates interactions of eIF3 and eIF2 on the ribosome required to position the initiator tRNA on the mRNA in the ribosomal peptidyl-tRNA site (P site).
Figures
Comment in
-
Biography of Jennifer A. Doudna.Proc Natl Acad Sci U S A. 2004 Dec 7;101(49):16987-9. doi: 10.1073/pnas.0408147101. Epub 2004 Dec 1. Proc Natl Acad Sci U S A. 2004. PMID: 15574498 Free PMC article. No abstract available.
References
-
- Hershey, J. W. B. & Merrick, W. C. (2000) in Translational Control of Gene Expression, eds. Sonenberg, N., Hershey, J. W. B. & Mathews, M. B. (Cold Spring Harbor Lab. Press, Plainview, NY), Vol. 39, pp. 33–88.
-
- Hellen, C. U. & Sarnow, P. (2001) Genes Dev. 15, 1593–1612. - PubMed
-
- Merrick, W. C. (2004) Gene 332, 1–11. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
