

Loss of function of KRE5 suppresses temperature sensitivity of mutants lacking mitochondrial anionic lipids
- PMID: 15563612
- PMCID: PMC545902
- DOI: 10.1091/mbc.e04-09-0808
Loss of function of KRE5 suppresses temperature sensitivity of mutants lacking mitochondrial anionic lipids
Erratum in
- Mol Biol Cell. 2005 Mar;16(3):1568
Abstract
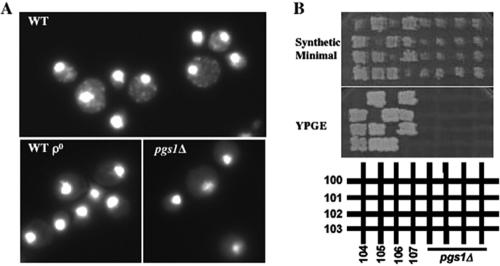
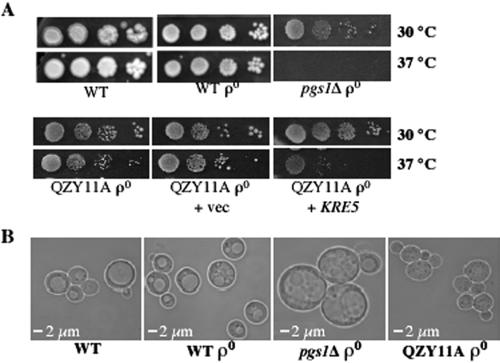
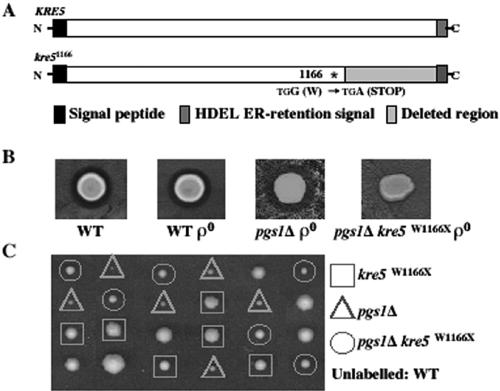
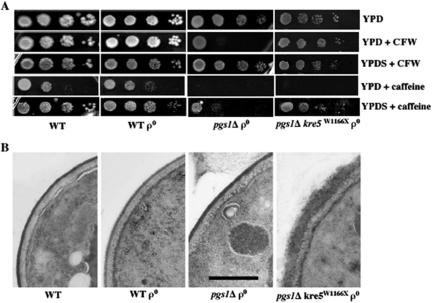
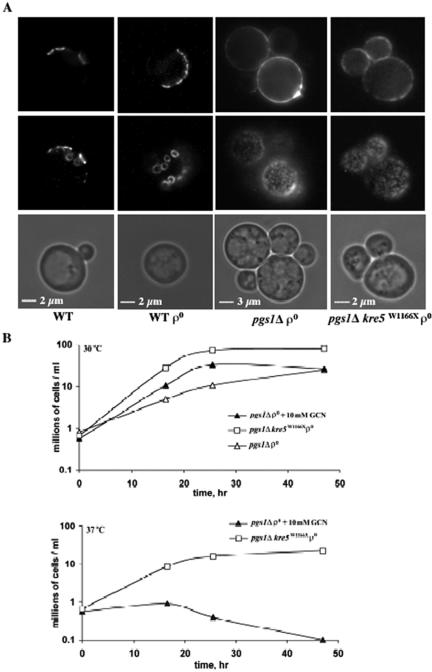
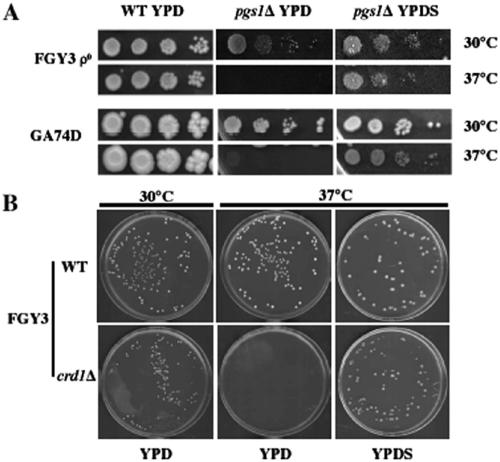
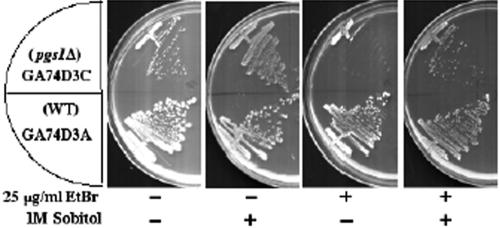
Disruption of PGS1, which encodes the enzyme that catalyzes the committed step of cardiolipin (CL) synthesis, results in loss of the mitochondrial anionic phospholipids phosphatidylglycerol (PG) and CL. The pgs1Delta mutant exhibits severe growth defects at 37 degrees C. To understand the essential functions of mitochondrial anionic lipids at elevated temperatures, we isolated suppressors of pgs1Delta that grew at 37 degrees C. One of the suppressors has a loss of function mutation in KRE5, which is involved in cell wall biogenesis. The cell wall of pgs1Delta contained markedly reduced beta-1,3-glucan, which was restored in the suppressor. Stabilization of the cell wall with osmotic support alleviated the cell wall defects of pgs1Delta and suppressed the temperature sensitivity of all CL-deficient mutants. Evidence is presented suggesting that the previously reported inability of pgs1Delta to grow in the presence of ethidium bromide was due to defective cell wall integrity, not from "petite lethality." These findings demonstrated that mitochondrial anionic lipids are required for cellular functions that are essential in cell wall biogenesis, the maintenance of cell integrity, and survival at elevated temperature.
Figures
References
-
- Alonso-Monge, R., Real, E., Wojda, I., Bebelman, J. P., Mager, W. H., and Siderius, M. (2001). Hyperosmotic stress response and regulation of cell wall integrity in Saccharomyces cerevisiae share common functional aspects. Mol. Microbiol. 41, 717-730. - PubMed
-
- Attfield, P. V. (1987). Trehalose accumulates in Saccharomyces cerevisiae during exposure to agents that induce heat shock response. FEBS Lett. 225, 259-263. - PubMed
-
- Barth, P. G., Scholte, H. R., Berden, J. A., Van der Klei-Van Moorsel, J. M., Luyt-Houwen, I. E., Van 't Veer-Korthof, E. T., Van der Harten, J. J., and Sobotka-Plojhar, M. A. (1983). An X-linked mitochondrial disease affecting cardiac muscle, skeletal muscle and neutrophil leucocytes. J. Neurol. Sci. 62, 327-355. - PubMed
-
- Barth, P. G., Van den Bogert, C., Bolhuis, P. A., Scholte, H. R., van Gennip, A. H., Schutgens, R. B., and Ketel, A. G. (1996). X-linked cardioskeletal myopathy and neutropenia (Barth syndrome): respiratory-chain abnormalities in cultured fibroblasts. J. Inherit. Metab. Dis. 19, 157-160. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
