

Protein transduction domains fused to virus receptors improve cellular virus uptake and enhance oncolysis by tumor-specific replicating vectors
- PMID: 15564483
- PMCID: PMC533898
- DOI: 10.1128/JVI.78.24.13743-13754.2004
Protein transduction domains fused to virus receptors improve cellular virus uptake and enhance oncolysis by tumor-specific replicating vectors
Abstract
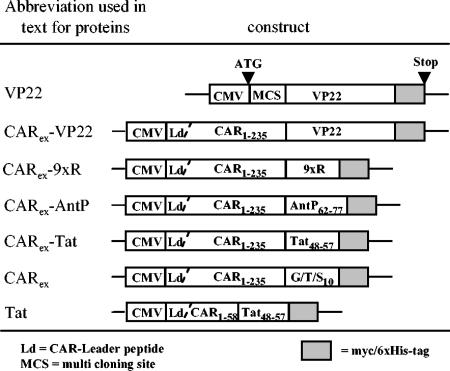
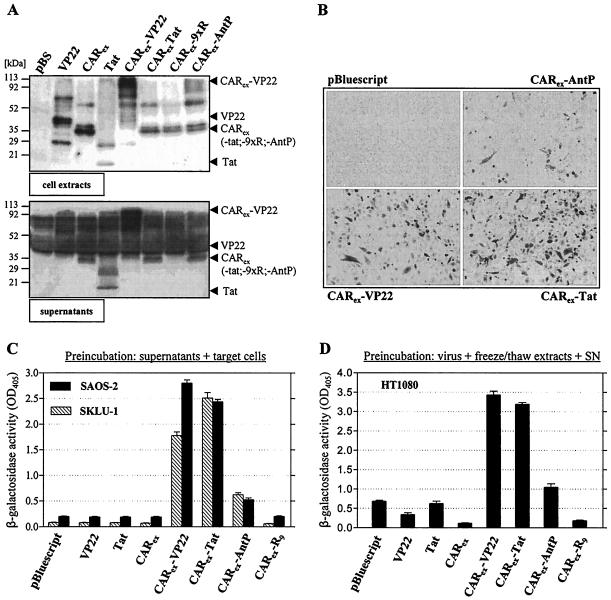
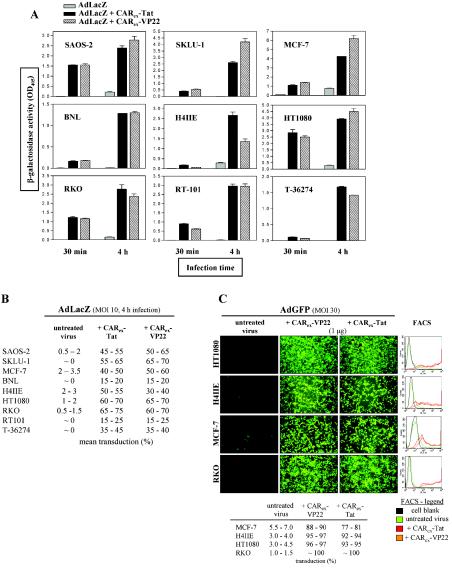
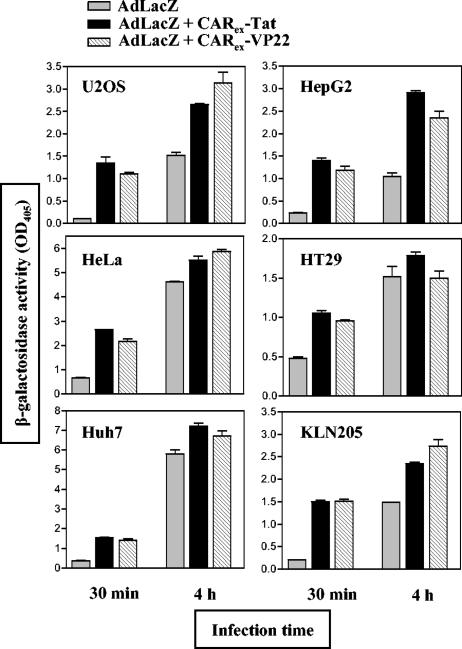
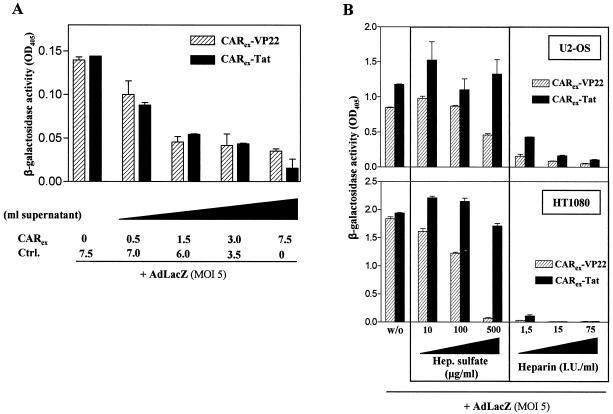
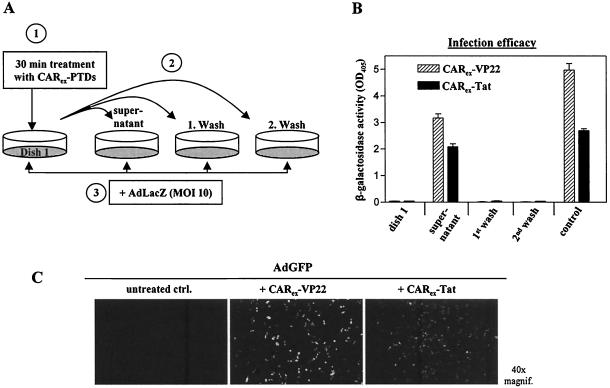
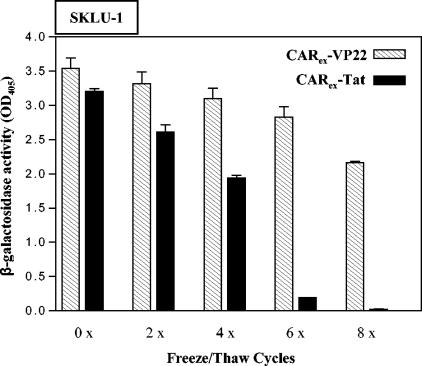
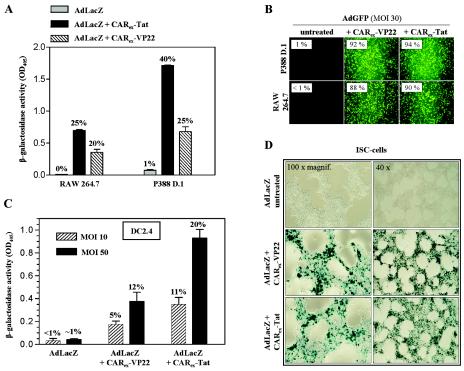
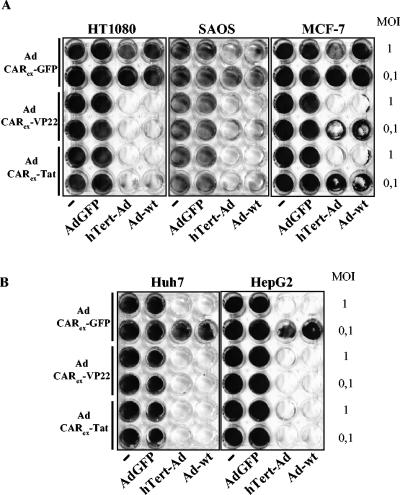
Expression of cellular receptors determines viral tropism and limits gene delivery by viral vectors. Protein transduction domains (PTDs) have been shown to deliver proteins, antisense oligonucleotides, liposomes, or plasmid DNA into cells. In our study, we investigated the role of several PTD motifs in adenoviral infection. When physiologically expressed, a PTD from human immunodeficiency virus transactivator of transcription (Tat) did not improve adenoviral infection. We therefore fused PTDs to the ectodomain of the coxsackievirus-adenovirus receptor (CAR(ex)) to attach PTDs to adenoviral fiber knobs. CAR(ex)-Tat and CAR(ex)-VP22 allowed efficient adenoviral infection in nonpermissive cells and significantly improved viral uptake rates in permissive cells. Dose-dependent competition of CAR(ex)-PTD-mediated infection using CAR(ex) and inhibition experiments with heparin showed that binding of CAR(ex)-PTD to both adenoviral fiber and cellular glycosaminoglycans is essential for the improvement of infection. CAR(ex)-PTD-treated adenoviruses retained their properties after density gradient ultracentrifugation, indicating stable binding of CAR(ex)-PTD to adenoviral particles. Consequently, the mechanism of CAR(ex)-PTD-mediated infection involves coating of the viral fiber knobs by CAR(ex)-PTD, rather than placement of CAR(ex) domains on cell surfaces. Expression of CAR(ex)-PTDs led to enhanced lysis of permissive and nonpermissive tumor cells by replicating adenoviruses, indicating that CAR(ex)-PTDs are valuable tools to improve the efficacy of oncolytic therapy. Together, our study shows that CAR(ex)-PTDs facilitate gene transfer in nonpermissive cells and improve viral uptake at reduced titers and infection times. The data suggest that PTDs fused to virus binding receptors may be a valuable tool to overcome natural tropism of vectors and could be of great interest for gene therapeutic approaches.
Figures
Similar articles
-
Efficient co-transduction of adenoviral vectors encoding carcinoembryonic antigen and survivin into dendritic cells by the CAR-TAT adaptor molecule enhance anti-tumor immunity in a murine colorectal cancer model.Immunol Lett. 2010 Jun 15;131(1):73-80. doi: 10.1016/j.imlet.2010.03.001. Epub 2010 Mar 6. Immunol Lett. 2010. PMID: 20211203
-
Fiber-modified adenovirus vectors containing the TAT peptide derived from HIV-1 in the fiber knob have efficient gene transfer activity.Gene Ther. 2007 Aug;14(15):1160-5. doi: 10.1038/sj.gt.3302969. Epub 2007 May 17. Gene Ther. 2007. PMID: 17508008
-
Tat conjugation of adenovirus vector broadens tropism and enhances transduction efficiency.Life Sci. 2008 Nov 21;83(21-22):747-55. doi: 10.1016/j.lfs.2008.09.022. Epub 2008 Oct 5. Life Sci. 2008. PMID: 18930744
-
Highly variable expression of virus receptors in the human cardiovascular system. Implications for cardiotropic viral infections and gene therapy.Z Kardiol. 2002 Dec;91(12):978-91. doi: 10.1007/s00392-002-0862-7. Z Kardiol. 2002. PMID: 12490988 Review.
-
Modified adenoviruses for cancer gene therapy.Int J Cancer. 2004 Jul 1;110(4):475-80. doi: 10.1002/ijc.20129. Int J Cancer. 2004. PMID: 15122578 Review.
Cited by
-
The Utilization of Cell-Penetrating Peptides in the Intracellular Delivery of Viral Nanoparticles.Materials (Basel). 2019 Aug 22;12(17):2671. doi: 10.3390/ma12172671. Materials (Basel). 2019. PMID: 31443361 Free PMC article. Review.
-
p53-dependent antiviral RNA-interference facilitates tumor-selective viral replication.Nucleic Acids Res. 2009 Jul;37(12):e84. doi: 10.1093/nar/gkp374. Epub 2009 May 14. Nucleic Acids Res. 2009. PMID: 19443444 Free PMC article.
-
Mesenchymal stem cells as carriers and amplifiers in CRAd delivery to tumors.Mol Cancer. 2011 Nov 3;10:134. doi: 10.1186/1476-4598-10-134. Mol Cancer. 2011. PMID: 22054049 Free PMC article.
-
Genetic incorporation of the protein transduction domain of Tat into Ad5 fiber enhances gene transfer efficacy.Virol J. 2007 Oct 24;4:103. doi: 10.1186/1743-422X-4-103. Virol J. 2007. PMID: 17958897 Free PMC article.
-
Selectivity of oncolytic viral replication prevents antiviral immune response and toxicity, but does not improve antitumoral immunity.Mol Ther. 2010 Nov;18(11):1972-82. doi: 10.1038/mt.2010.163. Epub 2010 Aug 10. Mol Ther. 2010. PMID: 20700112 Free PMC article.
References
-
- Bergelson, J. M., J. A. Cunningham, G. Droguett, E. A. Kurt-Jones, A. Krithivas, J. S. Hong, M. S. Horwitz, R. L. Crowell, and R. W. Finberg. 1997. Isolation of a common receptor for coxsackie B viruses and adenoviruses 2 and 5. Science 275:1320-1323. - PubMed
-
- Console, S., C. Marty, C. Garcia-Echeverria, R. Schwendener, and K. Ballmer-Hofer. 2003. Antennapedia and HIV transactivator of transcription (TAT) “protein transduction domains” promote endocytosis of high molecular weight cargo upon binding to cell surface glycosaminoglycans. J. Biol. Chem. 278:35109-35114. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
