

Targeted disruption of luteinizing hormone beta-subunit leads to hypogonadism, defects in gonadal steroidogenesis, and infertility
- PMID: 15569941
- PMCID: PMC535369
- DOI: 10.1073/pnas.0404743101
Targeted disruption of luteinizing hormone beta-subunit leads to hypogonadism, defects in gonadal steroidogenesis, and infertility
Abstract
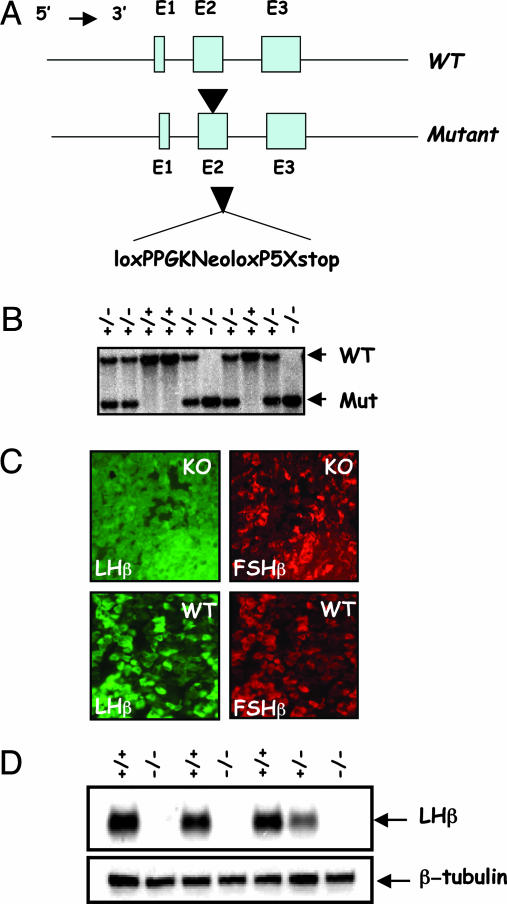
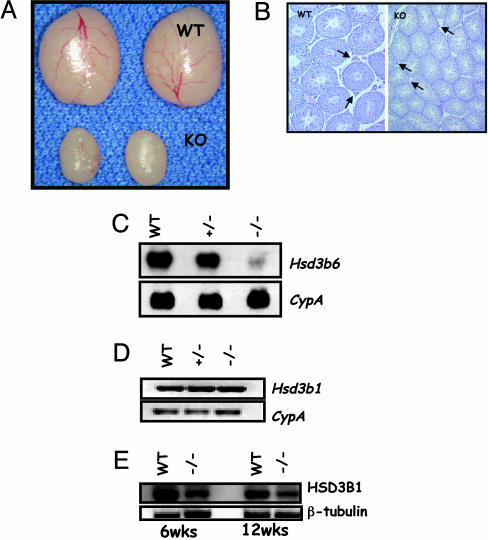
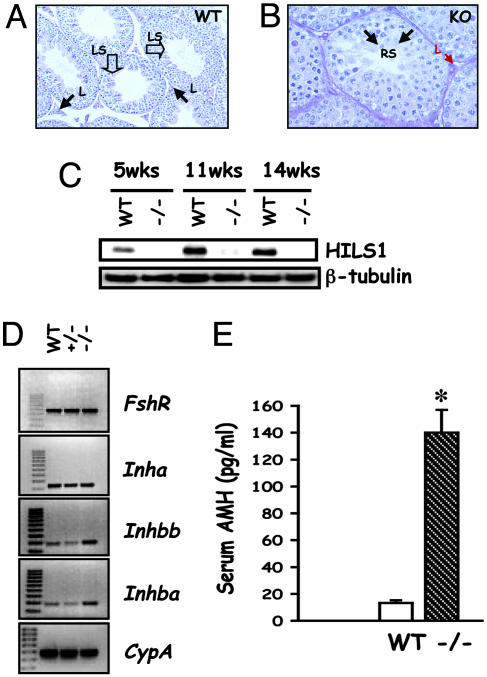
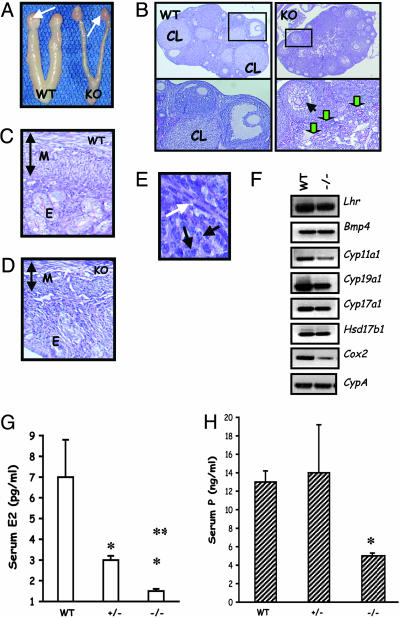
Luteinizing hormone (LH) and follicle-stimulating hormone (FSH) act on gonadal cells to promote steroidogenesis and gametogenesis. Clarifying the in vivo roles of LH and FSH permits a feasible approach to contraception involving selective blockade of gonadotropin action. One way to address these physiologically important problems is to generate mice with an isolated LH deficiency and compare them with existing FSH loss-of-function mice. To model human reproductive disorders involving loss of LH function and to define LH-responsive genes, we produced knockout mice lacking the hormone-specific LHbeta-subunit. LHbeta-null mice are viable but demonstrate postnatal defects in gonadal growth and function resulting in infertility. Mutant males have decreased testes size, prominent Leydig cell hypoplasia, defects in expression of genes encoding steroid biosynthesis pathway enzymes, and reduced testosterone levels. Furthermore, spermatogenesis is blocked at the round spermatid stage, causing a total absence of the elongated spermatids. Mutant female mice are hypogonadal and demonstrate decreased levels of serum estradiol and progesterone. Ovarian histology demonstrates normal thecal layer, defects in folliculogenesis including many degenerating antral follicles, and absence of corpora lutea. The defects in both sexes are not secondary to aberrant FSH regulation, because FSH levels were unaffected in null mice. Finally, both male and female null mice can be pharmacologically rescued by exogenous human chorionic gonadotropin, indicating that LH-responsiveness of the target cells is not irreversibly lost. Thus, LHbeta null mice represent a model to study the consequences of an isolated deficiency of LH ligand in reproduction, while retaining normal LH-responsiveness in target cells.
Figures
References
-
- Pierce, J. G. & Parsons, T. F. (1981) Annu. Rev. Biochem. 50, 465–495. - PubMed
-
- Bousfield, G. R., Perry, W. M. & Ward, D. N. (1994) in The Physiology of Reproduction, eds. Knobil, E. & Neill, J. D. (Raven, New York), pp. 1749–1792.
-
- O'Shaughnessy, P. J., Baker, P., Sohnius, U., Haavisto, A. M., Charlton, H. M. & Huhtaniemi, I. (1998) Endocrinology 139, 1141–1146. - PubMed
-
- Pakarinen, P., Kimura, S., El-Gehani, F., Pelliniemi, L. J. & Huhtaniemi, I. (2002) Endocrinology 143, 4477–4482. - PubMed
-
- Saez, J. M., Perrard-Sapori, M. H., Chatelain, P. G., Tabone, E. & Rivarola, M. A. (1987) J. Steroid. Biochem. 27, 317–329. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
