

Role of connexin-based gap junction channels and hemichannels in ischemia-induced cell death in nervous tissue
- PMID: 15572178
- PMCID: PMC3651737
- DOI: 10.1016/j.brainresrev.2004.08.002
Role of connexin-based gap junction channels and hemichannels in ischemia-induced cell death in nervous tissue
Abstract
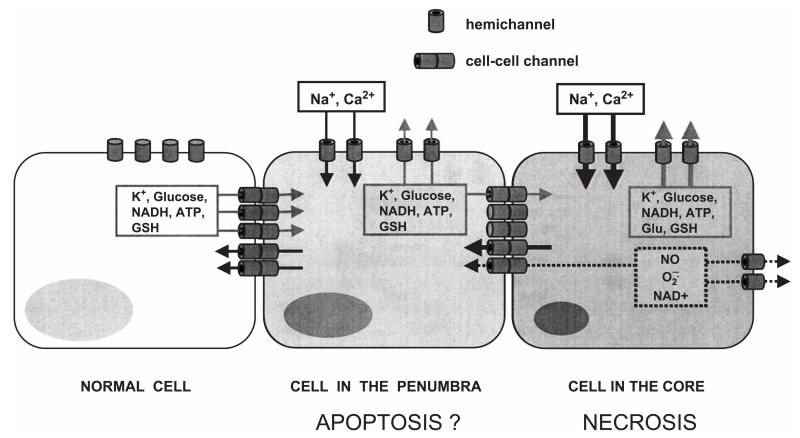
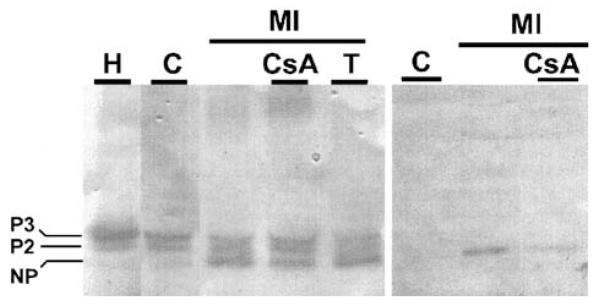
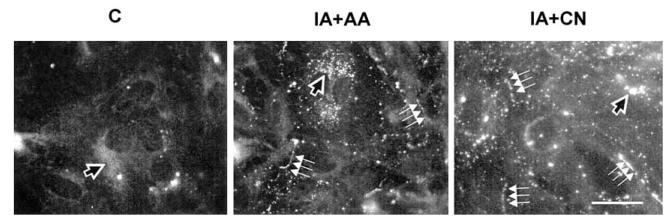
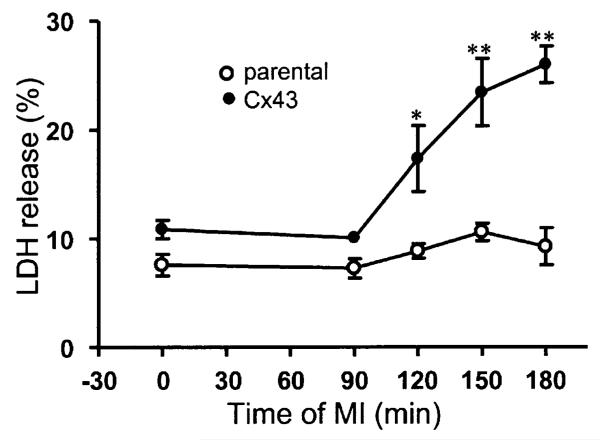
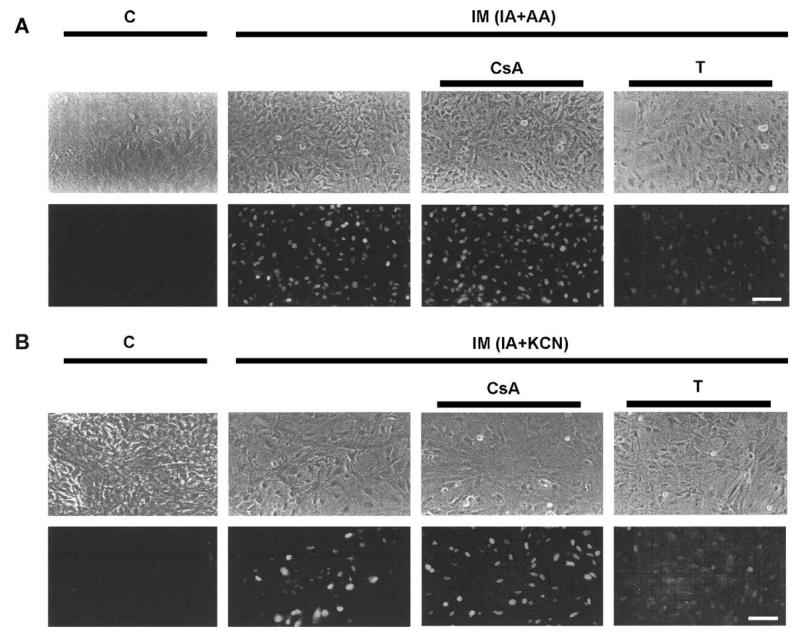
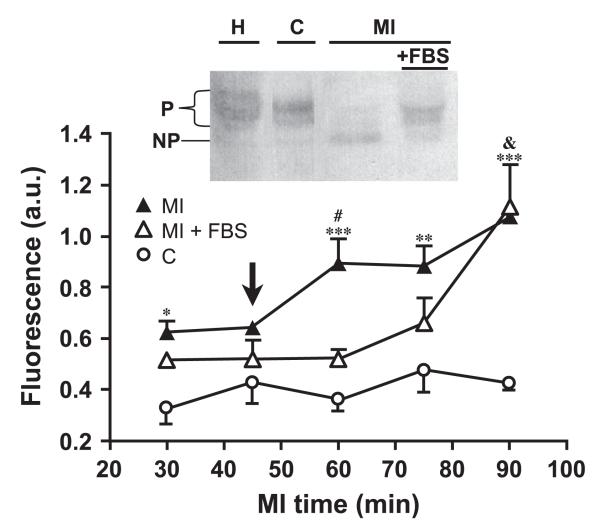
Gap junction channels and hemichannels formed of connexin subunits are found in most cell types in vertebrates. Gap junctions connect cells via channels not open to the extracellular space and permit the passage of ions and molecules of approximately 1 kDa. Single connexin hemichannels, which are connexin hexamers, are present in the surface membrane before docking with a hemichannel in an apposed membrane. Because of their high conductance and permeability in cell-cell channels, it had been thought that connexin hemichannels remained closed until docking to form a cell-cell channel. Now it is clear that at least some hemichannels can open to allow passage of molecules between the cytoplasm and extracellular space. Here we review evidence that gap junction channels may allow intercellular diffusion of necrotic or apoptotic signals, but may also allow diffusion of ions and substances from healthy to injured cells, thereby contributing to cell survival. Moreover, opening of gap junction hemichannels may exacerbate cell injury or mediate paracrine or autocrine signaling. In addition to the cell specific features of an ischemic insult, propagation of cell damage and death within affected tissues may be affected by expression and regulation of gap junction channels and hemichannels formed by connexins.
Figures
Similar articles
-
Hemichannels in cerebral ischemia.Curr Mol Med. 2009 Mar;9(2):186-94. doi: 10.2174/156652409787581646. Curr Mol Med. 2009. PMID: 19275626 Review.
-
Emerging issues of connexin channels: biophysics fills the gap.Q Rev Biophys. 2001 Aug;34(3):325-472. doi: 10.1017/s0033583501003705. Q Rev Biophys. 2001. PMID: 11838236 Review.
-
Ischemia induces closure of gap junctional channels and opening of hemichannels in heart-derived cells and tissue.Cell Physiol Biochem. 2011;28(1):103-14. doi: 10.1159/000331719. Epub 2011 Aug 16. Cell Physiol Biochem. 2011. PMID: 21865853
-
Regulation of gap junction channels and hemichannels by phosphorylation and redox changes: a revision.BMC Cell Biol. 2016 May 24;17 Suppl 1(Suppl 1):11. doi: 10.1186/s12860-016-0099-3. BMC Cell Biol. 2016. PMID: 27229925 Free PMC article. Review.
-
Connexin-based gap junction hemichannels: gating mechanisms.Biochim Biophys Acta. 2005 Jun 10;1711(2):215-24. doi: 10.1016/j.bbamem.2005.01.014. Epub 2005 Mar 2. Biochim Biophys Acta. 2005. PMID: 15955306 Free PMC article. Review.
Cited by
-
Further evidence for P59L mutation in GJA3 associated with autosomal dominant congenital cataract.Indian J Ophthalmol. 2016 Jul;64(7):508-12. doi: 10.4103/0301-4738.190139. Indian J Ophthalmol. 2016. PMID: 27609163 Free PMC article.
-
The Aerial Parts of Bupleurum Chinense DC. Aromatic Oil Attenuate Kainic Acid-Induced Epilepsy-Like Behavior and Its Potential Mechanisms.Biomed Res Int. 2022 Apr 11;2022:1234612. doi: 10.1155/2022/1234612. eCollection 2022. Biomed Res Int. 2022. PMID: 35445130 Free PMC article.
-
Modulation of Cx46 hemichannels by nitric oxide.Am J Physiol Cell Physiol. 2009 Jun;296(6):C1356-63. doi: 10.1152/ajpcell.00054.2009. Epub 2009 Apr 8. Am J Physiol Cell Physiol. 2009. PMID: 19357237 Free PMC article.
-
Cardiac to cancer: connecting connexins to clinical opportunity.FEBS Lett. 2014 Apr 17;588(8):1349-64. doi: 10.1016/j.febslet.2014.02.047. Epub 2014 Mar 4. FEBS Lett. 2014. PMID: 24607540 Free PMC article. Review.
-
Disruption of ion homeostasis in the neurogliovascular unit underlies the pathogenesis of ischemic cerebral edema.Transl Stroke Res. 2014 Feb;5(1):3-16. doi: 10.1007/s12975-013-0307-9. Epub 2013 Nov 22. Transl Stroke Res. 2014. PMID: 24323726 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
