

Identification of a unique filamin A binding region within the cytoplasmic domain of glycoprotein Ibalpha
- PMID: 15574123
- PMCID: PMC1135017
- DOI: 10.1042/BJ20041836
Identification of a unique filamin A binding region within the cytoplasmic domain of glycoprotein Ibalpha
Abstract
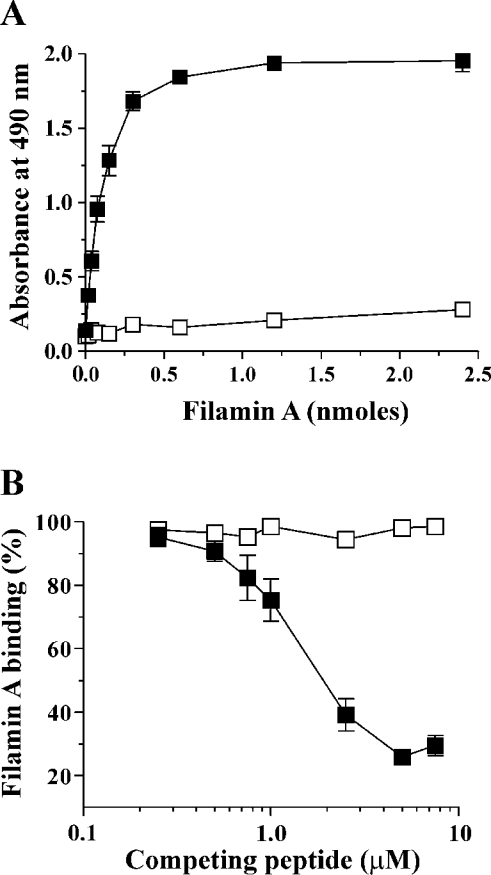
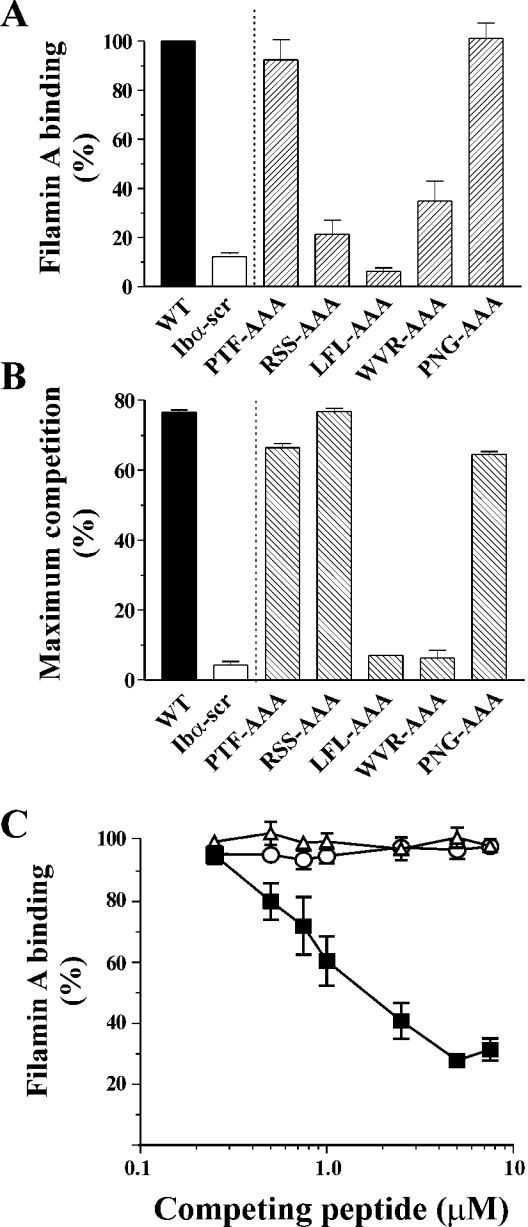
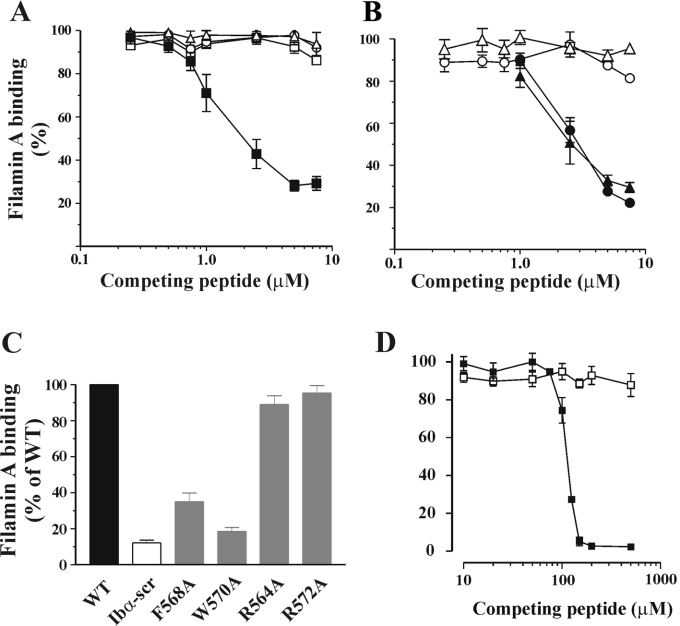
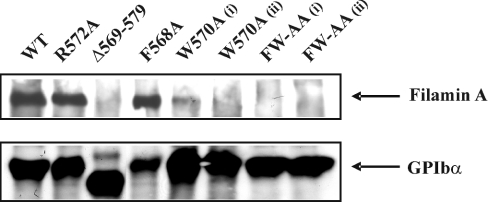
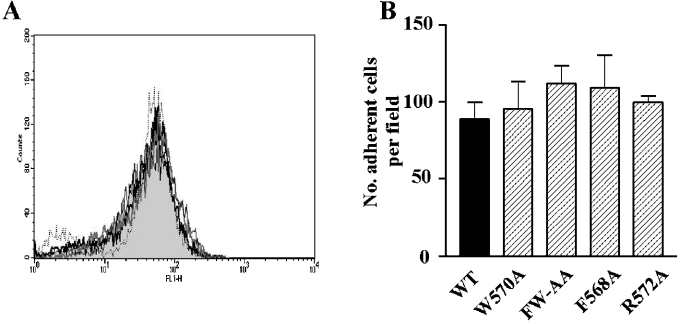
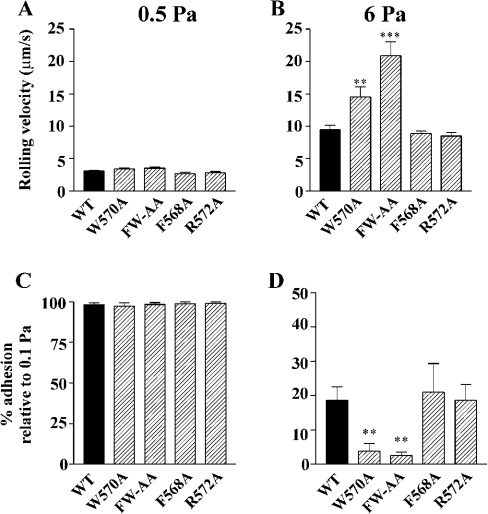
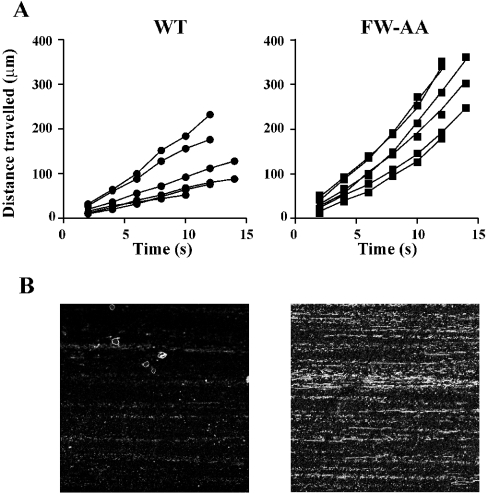
Binding of the platelet GPIb/V/IX (glycoprotein Ib/V/IX) receptor to von Willebrand factor is critical for platelet adhesion and aggregation under conditions of rapid blood flow. The adhesive function of GPIbalpha is regulated by its anchorage to the membrane skeleton through a specific interaction with filamin A. In the present study, we examined the amino acid residues within the cytoplasmic tail of GPIbalpha, which are critical for association with filamin A, using a series of 25-mer synthetic peptides that mimic the cytoplasmic tail sequences of wild-type and mutant forms of GPIbalpha. Peptide binding studies of purified human filamin A have demonstrated a major role for the conserved hydrophobic stretch L567FLWV571 in mediating this interaction. Progressive alanine substitutions of triple, double and single amino acid residues within the Pro561-Arg572 region suggested an important role for Trp570 and Phe568 in promoting GPIbalpha binding to filamin A. The importance of these two residues in promoting filamin A binding to GPIbalpha in vivo was confirmed from the study of Chinese-hamster ovary cells expressing GPIbalpha Trp570-->Ala and Phe568-->Ala substitutions. Phenotypic analysis of these cell lines in flow-based adhesion studies revealed a critical role for these residues in maintaining receptor anchorage to the membrane skeleton and in maintaining cell adhesion to a von Willebrand factor matrix under high-shear conditions. These studies demonstrate a novel filamin A binding motif in the cytoplasmic tail of GPIbalpha, which is critically dependent on both Trp570 and Phe568.
Figures

Similar articles
-
Filamin A binding to the cytoplasmic tail of glycoprotein Ibalpha regulates von Willebrand factor-induced platelet activation.Blood. 2003 Sep 15;102(6):2122-9. doi: 10.1182/blood-2002-12-3805. Epub 2003 Jun 5. Blood. 2003. PMID: 12791664
-
Interaction between platelet glycoprotein Ibalpha and filamin-1 is essential for glycoprotein Ib/IX receptor anchorage at high shear.J Biol Chem. 2002 Jan 18;277(3):2151-9. doi: 10.1074/jbc.M109384200. Epub 2001 Nov 7. J Biol Chem. 2002. PMID: 11700320
-
Identification of a novel 14-3-3zeta binding site within the cytoplasmic domain of platelet glycoprotein Ibalpha that plays a key role in regulating the von Willebrand factor binding function of glycoprotein Ib-IX.Circ Res. 2009 Dec 4;105(12):1177-85. doi: 10.1161/CIRCRESAHA.109.204669. Epub 2009 Oct 29. Circ Res. 2009. PMID: 19875727
-
Functional association of phosphoinositide-3-kinase with platelet glycoprotein Ibalpha, the major ligand-binding subunit of the glycoprotein Ib-IX-V complex.J Thromb Haemost. 2010 Feb;8(2):324-30. doi: 10.1111/j.1538-7836.2009.03672.x. Epub 2009 Oct 30. J Thromb Haemost. 2010. PMID: 19874472
-
Signalling through the platelet glycoprotein Ib-V-IX complex.Cell Signal. 2004 Dec;16(12):1329-44. doi: 10.1016/j.cellsig.2004.05.008. Cell Signal. 2004. PMID: 15381249 Review.
Cited by
-
Isoform divergence of the filamin family of proteins.Mol Biol Evol. 2010 Feb;27(2):283-95. doi: 10.1093/molbev/msp236. Epub 2009 Oct 5. Mol Biol Evol. 2010. PMID: 19805437 Free PMC article.
-
Platelets and cancer: a casual or causal relationship: revisited.Cancer Metastasis Rev. 2014 Mar;33(1):231-69. doi: 10.1007/s10555-014-9498-0. Cancer Metastasis Rev. 2014. PMID: 24696047 Free PMC article. Review.
-
Membrane skeleton orchestrates the platelet glycoprotein (GP) Ib-IX complex clustering and signaling.IUBMB Life. 2016 Oct;68(10):823-9. doi: 10.1002/iub.1559. Epub 2016 Sep 15. IUBMB Life. 2016. PMID: 27634617 Free PMC article.
-
Platelet size matters.Blood. 2024 Jan 25;143(4):298-300. doi: 10.1182/blood.2023023057. Blood. 2024. PMID: 38270943 Free PMC article. No abstract available.
-
In Vitro Exposure to Glucose Alters the Expression of Phosphorylated Proteins in Platelets.Biomedicines. 2023 Feb 13;11(2):543. doi: 10.3390/biomedicines11020543. Biomedicines. 2023. PMID: 36831080 Free PMC article.
References
-
- Savage B., Saldivar E., Ruggeri Z. M. Initiation of platelet adhesion by arrest onto fibrinogen or translocation on von Willebrand factor. Cell (Cambridge, Mass.) 1996;84:289–297. - PubMed
-
- Berndt M. C., Shen Y., Dopheide S. M., Gardiner E. E., Andrews R. K. The vascular biology of the glycoprotein Ib-IX-V complex. Thromb. Haemostasis. 2001;86:178–188. - PubMed
-
- Lopez J. A., Andrews R. K., Afshar-Kharghan V., Berndt M. C. Bernard-Soulier syndrome. Blood. 1998;91:4397–4418. - PubMed
-
- Du X., Fox J. E., Pei S. Identification of a binding sequence for the 14-3-3 protein within the cytoplasmic domain of the adhesion receptor, platelet glycoprotein Ib alpha. J. Biol. Chem. 1996;271:7362–7367. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous