

Molecular force modulation spectroscopy revealing the dynamic response of single bacteriorhodopsins
- PMID: 15574708
- PMCID: PMC1305144
- DOI: 10.1529/biophysj.104.052746
Molecular force modulation spectroscopy revealing the dynamic response of single bacteriorhodopsins
Abstract
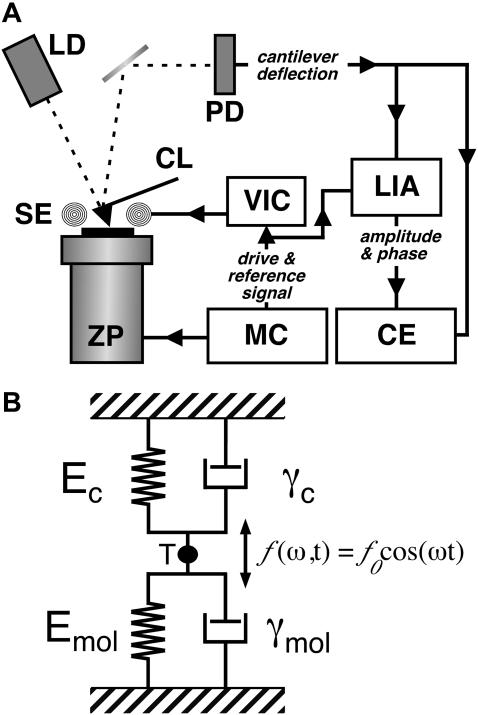
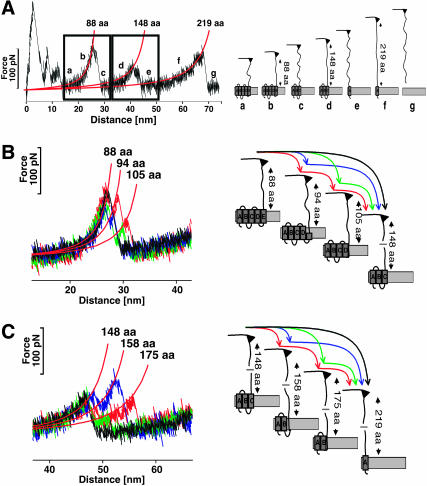
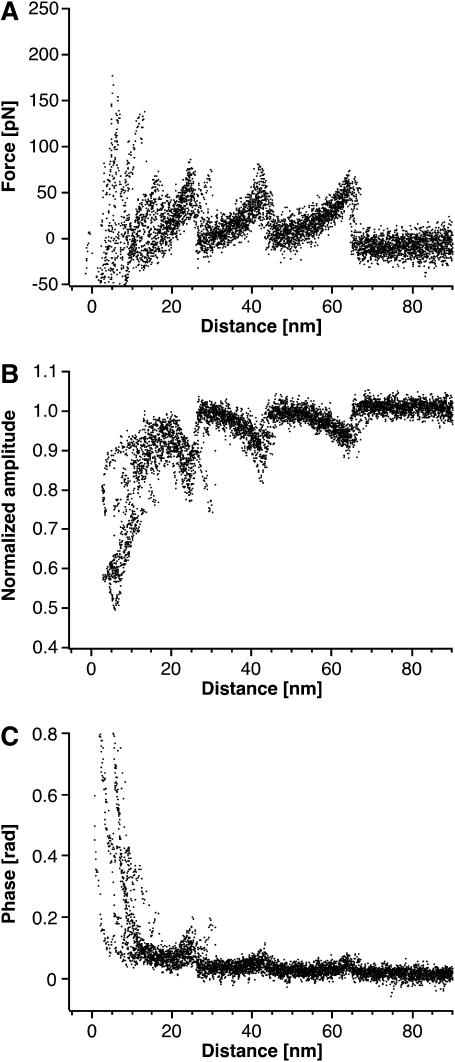
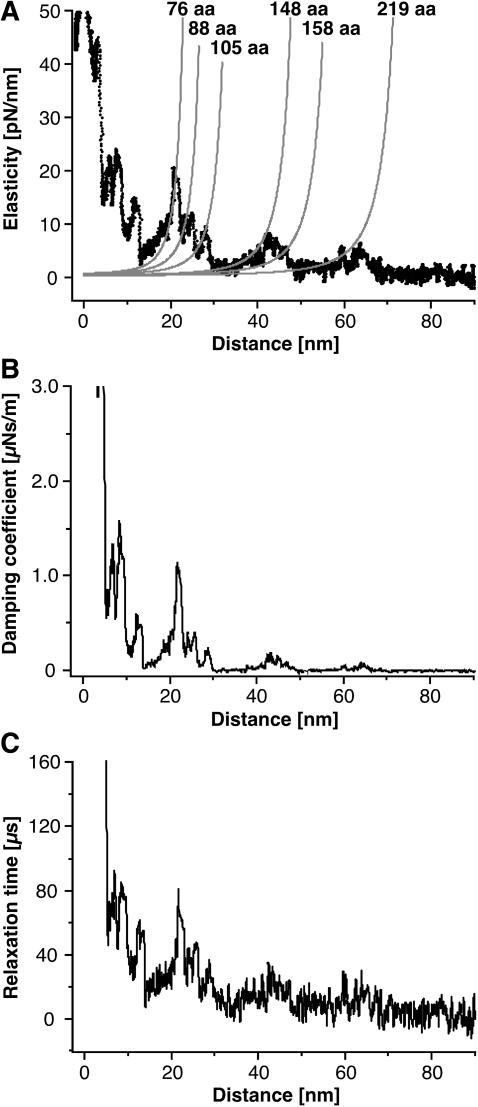
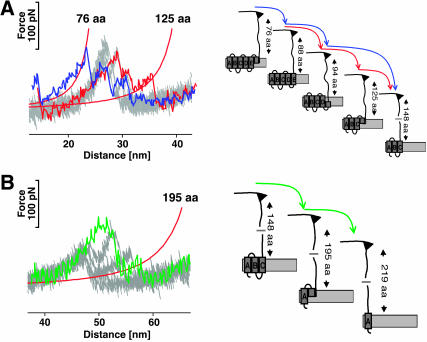
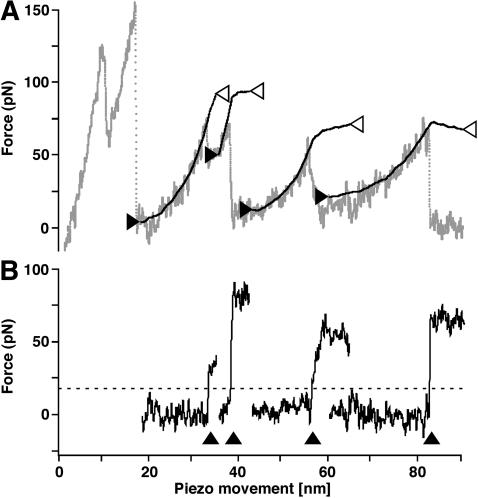
Recent advances in atomic force microscopy allowed globular and membrane proteins to be mechanically unfolded on a single-molecule level. Presented is an extension to the existing force spectroscopy experiments. While unfolding single bacteriorhodopsins from native purple membranes, small oscillation amplitudes (6-9 nm) were supplied to the vertical displacement of the cantilever at a frequency of 3 kHz. The phase and amplitude response of the cantilever-protein system was converted to reveal the elastic (conservative) and viscous (dissipative) contributions to the unfolding process. The elastic response (stiffness) of the extended parts of the protein were in the range of a few tens pN/nm and could be well described by the derivative of the wormlike chain model. Discrete events in the viscous response coincided with the unfolding of single secondary structure elements and were in the range of 1 microNs/m. In addition, these force modulation spectroscopy experiments revealed novel mechanical unfolding intermediates of bacteriorhodopsin. We found that kinks result in a loss of unfolding cooperativity in transmembrane helices. Reconstructing force-distance spectra by the integration of amplitude-distance spectra verified their position, offering a novel approach to detect intermediates during the forced unfolding of single proteins.
Figures
Comment in
-
Unraveling bacteriorhodopsin.Biophys J. 2005 Feb;88(2):763-4. doi: 10.1529/biophysj.104.056242. Epub 2004 Dec 13. Biophys J. 2005. PMID: 15596486 Free PMC article. No abstract available.
References
-
- Binnig, G., C. F. Quate, and C. Gerber. 1986. Atomic force microscope. Phys. Rev. Lett. 56:930–933. - PubMed
-
- Booth, P. J. 1997. Folding alpha-helical membrane proteins: kinetic studies on bacteriorhodopsin. Fold. Des. 2:R85–R92. - PubMed
-
- Booth, P. J., R. H. Templer, A. R. Curran, and S. J. Allen. 2001. Can we identify the forces that drive the folding of integral membrane proteins? Biochem. Soc. T. 29:408–413. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
