

Mapping netrin receptor binding reveals domains of Unc5 regulating its tyrosine phosphorylation
- PMID: 15574733
- PMCID: PMC6730211
- DOI: 10.1523/JNEUROSCI.3715-04.2004
Mapping netrin receptor binding reveals domains of Unc5 regulating its tyrosine phosphorylation
Abstract
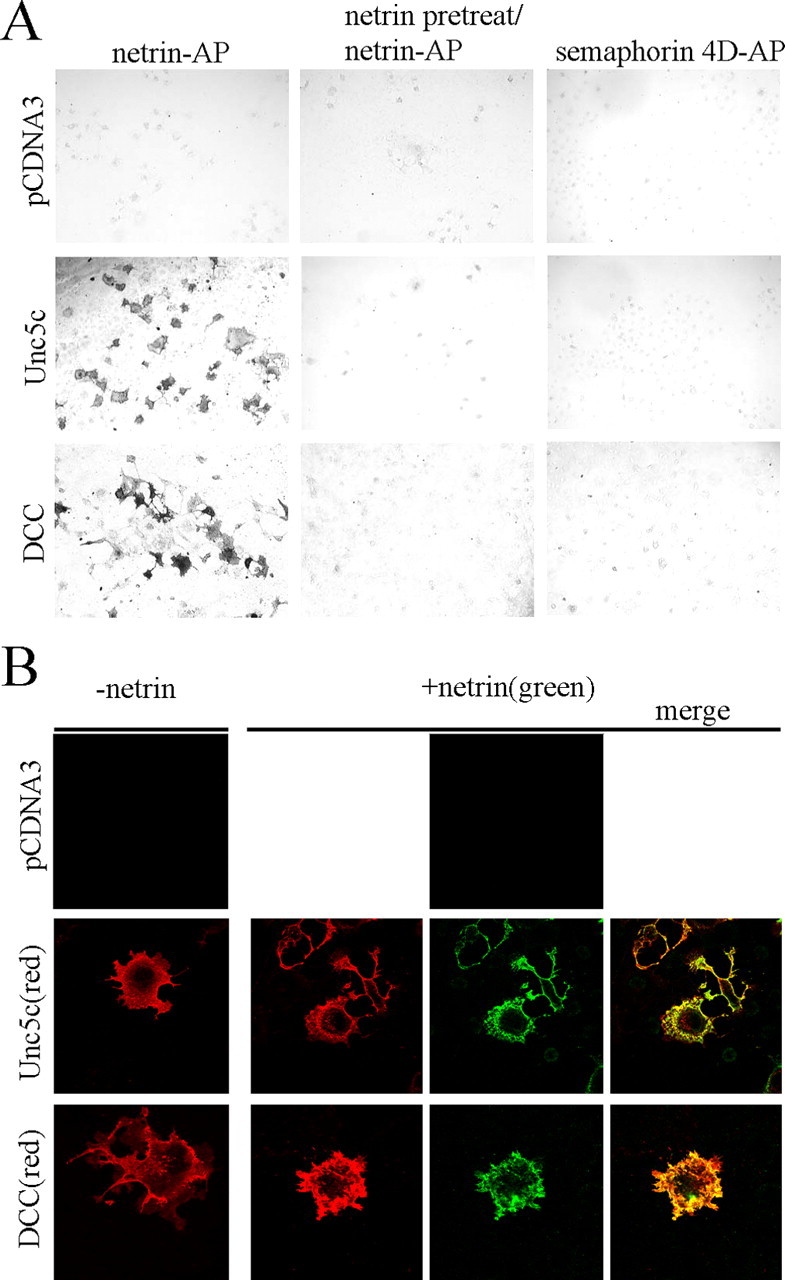
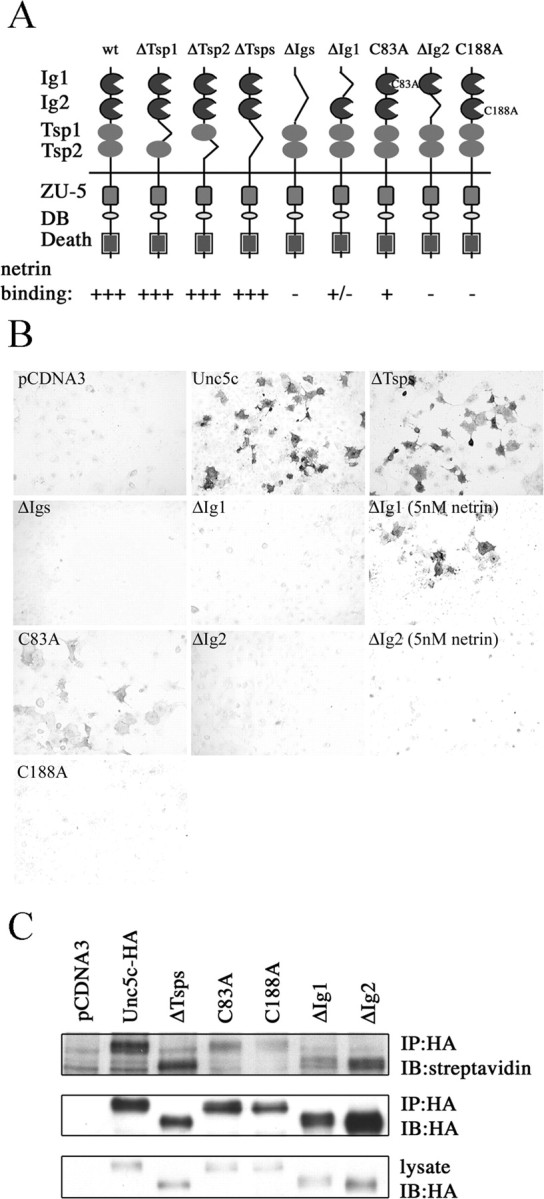
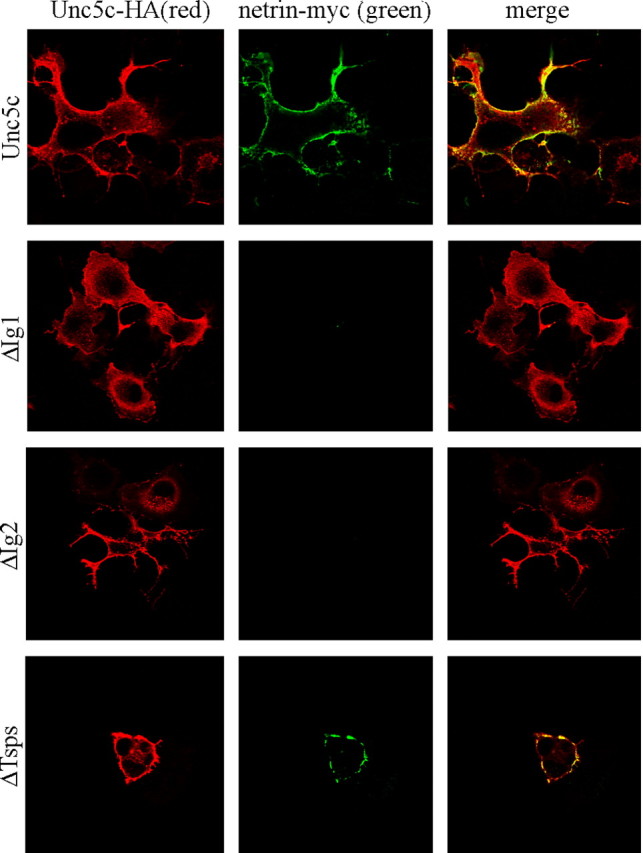
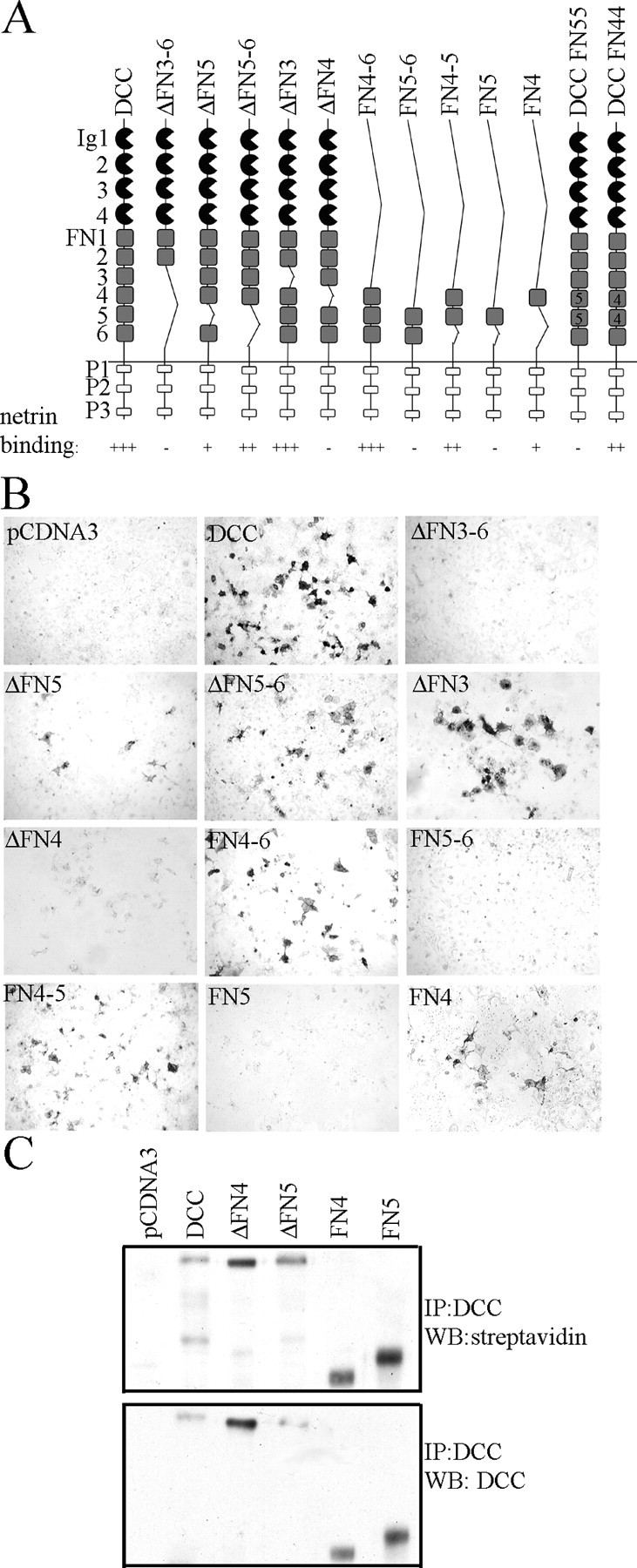
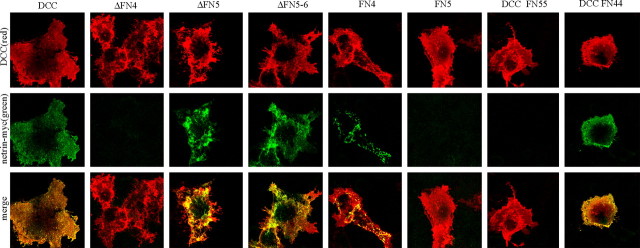
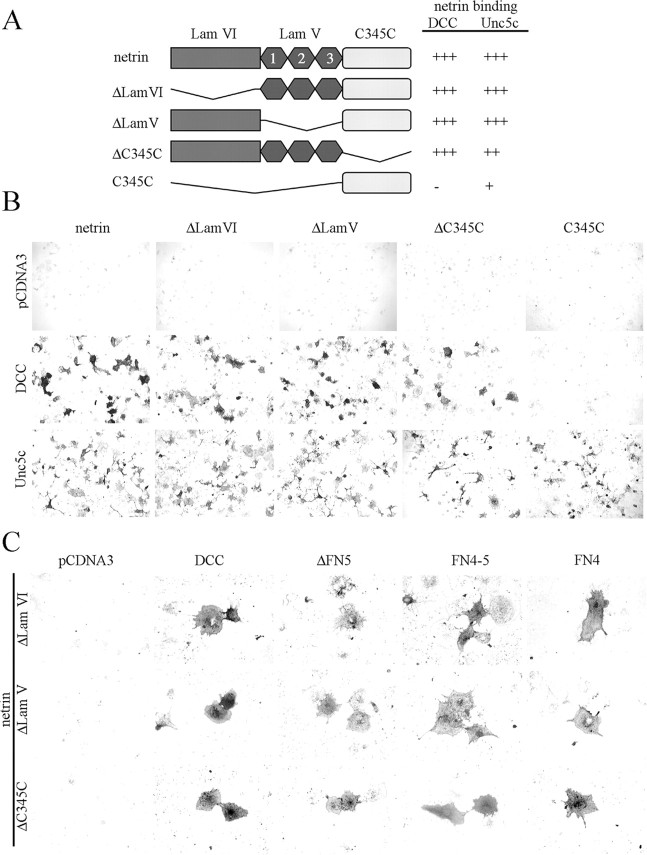
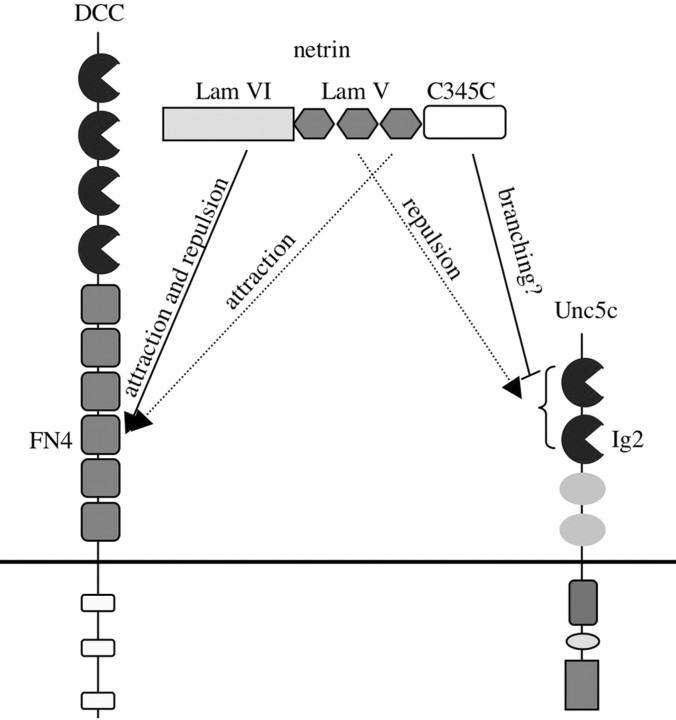
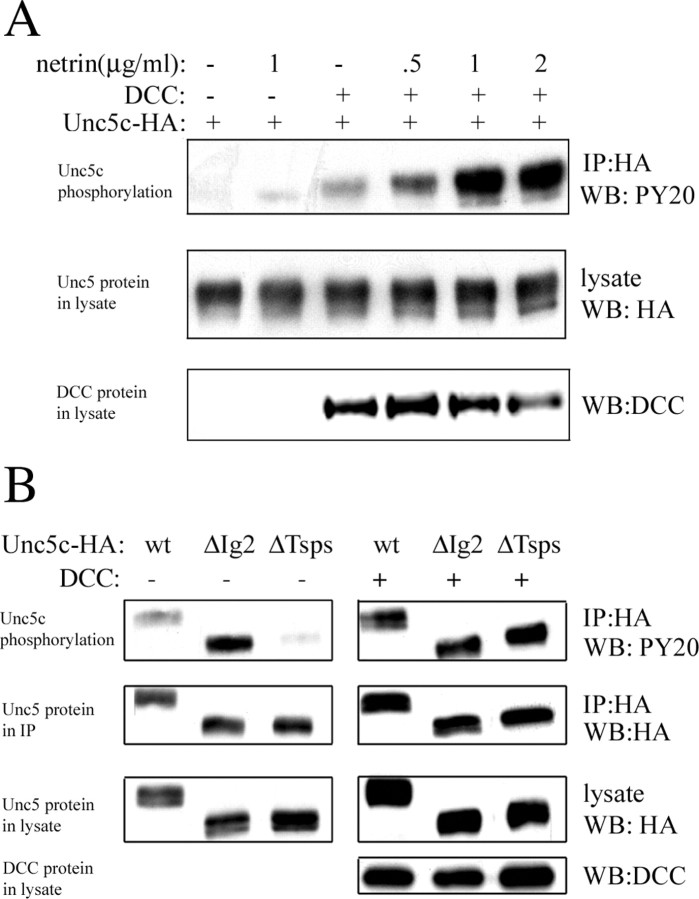
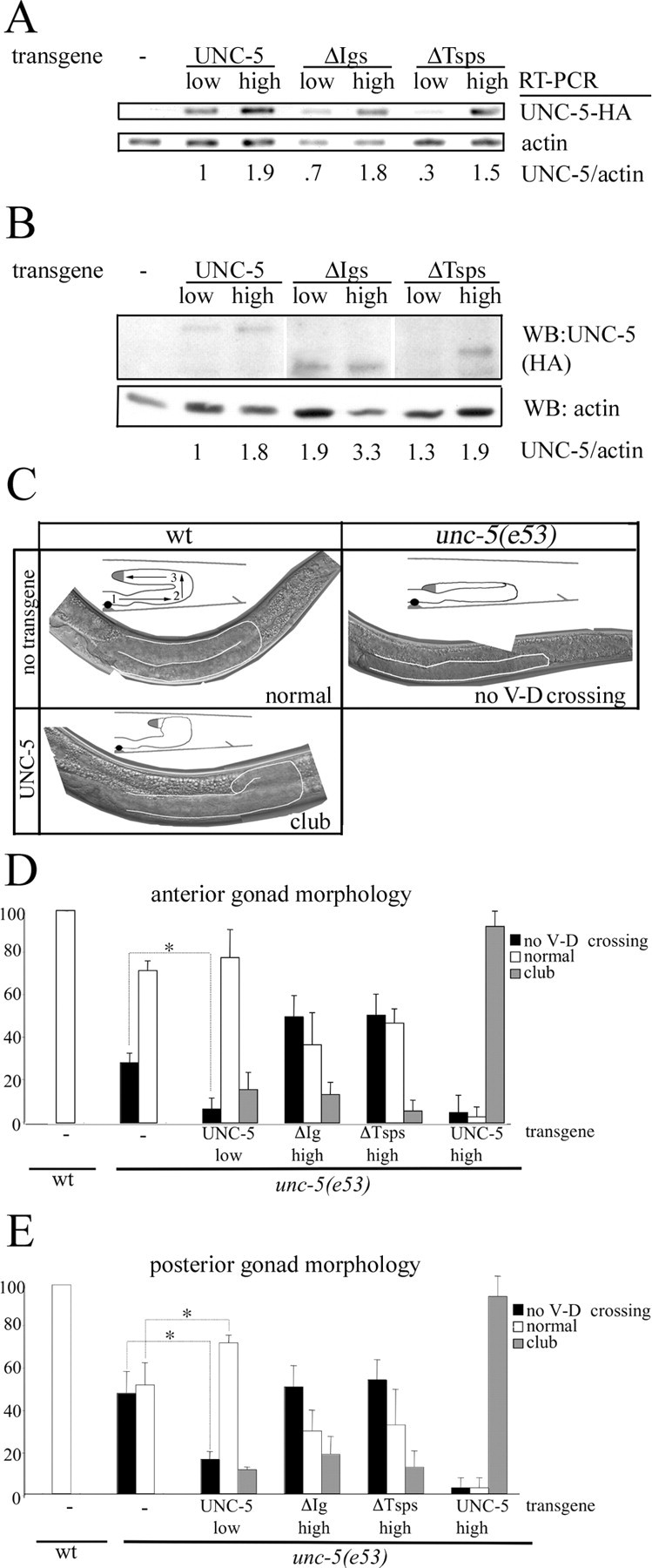
Netrin and its receptors Unc5 and deleted in colorectal carcinoma (DCC) regulate axon guidance and cell migration. We defined domains involved in the interactions between netrin-1, DCC, and Unc5c. We show that Unc5 requires both Ig domains to interact with netrin. DCC binds through the fourth fibronectin type III domain, whereas netrin binds through multiple domains to both receptors. We examined the functional consequences of removing the netrin binding and nonbinding domains from Unc5 in vitro and in vivo. In human embryonic kidney 293 cells, removal of the netrin binding second Ig domain causes an increase in basal tyrosine phosphorylation, whereas removal of the netrin nonbinding thrombospondin domains decreases tyrosine phosphorylation. Moreover, experiments in Caenorhabditis elegans indicate that both netrin binding and nonbinding domains are necessary for phenotypic rescue of an unc-5 loss of function mutation.
Figures
References
-
- Bennett KL, Bradshaw J, Youngman T, Rodgers J, Greenfield B, Aruffo A, Linsley PS (1997) Deleted in colorectal carcinoma (DCC) binds heparin via its fifth fibronectin type III domain. J Biol Chem 272: 26940-26946. - PubMed
-
- Chan SS, Zheng H, Su MW, Wilk R, Killeen MT, Hedgecock EM, Culotti JG (1996) UNC-40, a C. elegans homolog of DCC (Deleted in Colorectal Cancer), is required in motile cells responding to UNC-6 netrin cues. Cell 87: 187-195. - PubMed
-
- Colamarino SA, Tessier-Lavigne M (1995) The axonal chemoattractant netrin-1 is also a chemorepellent for trochlear motor axons. Cell 81: 621-629. - PubMed
-
- Colavita A, Krishna S, Zheng H, Padgett RW, Culotti JG (1998) Pioneer axon guidance by UNC-129, a C. elegans TGF-beta. Science 281: 706-709. - PubMed
-
- Dickson B (2002) Molecular mechanisms of axon guidance. Science 298: 1959-1964. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases