

Molecular basis for TPR domain-mediated regulation of protein phosphatase 5
- PMID: 15577939
- PMCID: PMC544909
- DOI: 10.1038/sj.emboj.7600496
Molecular basis for TPR domain-mediated regulation of protein phosphatase 5
Abstract
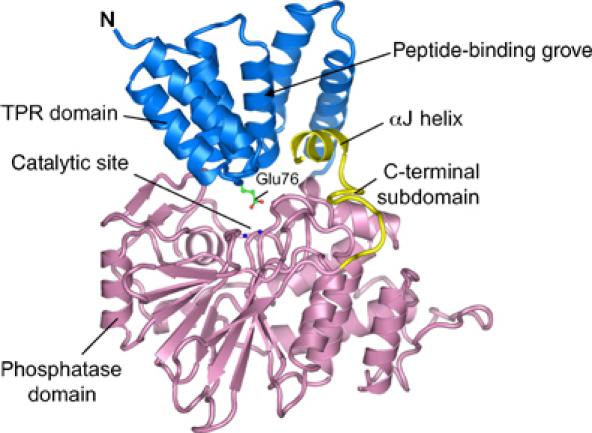
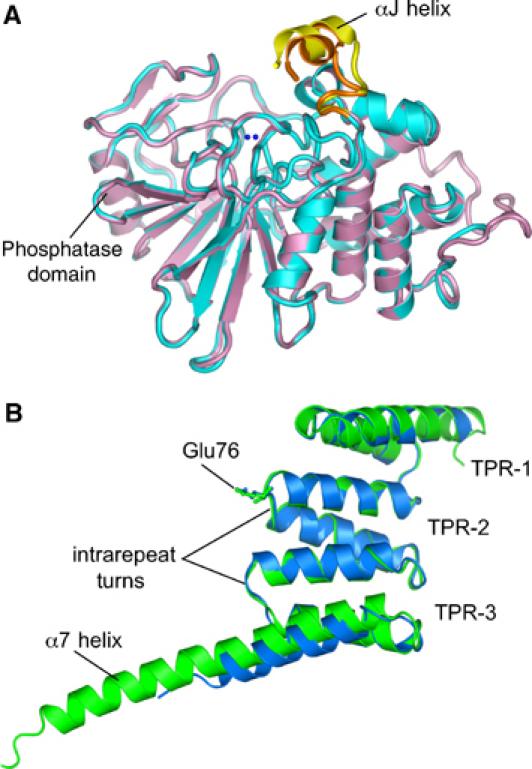
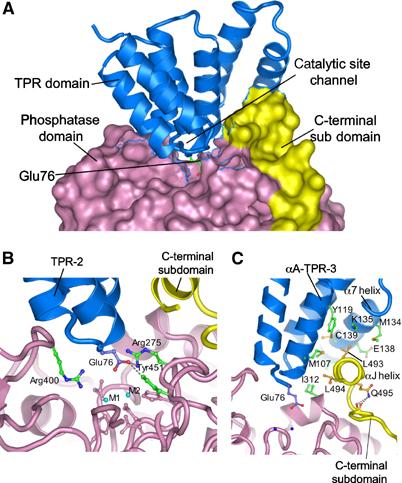
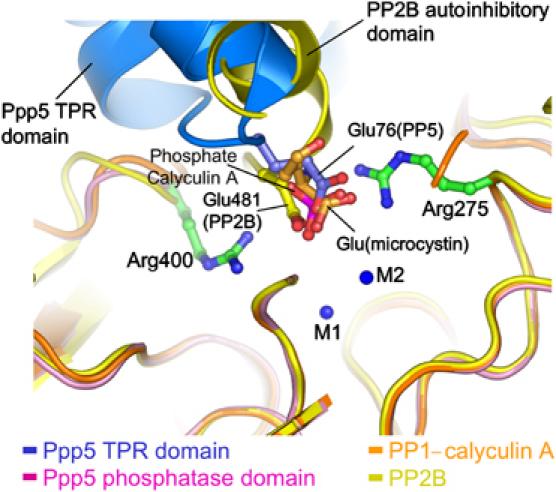
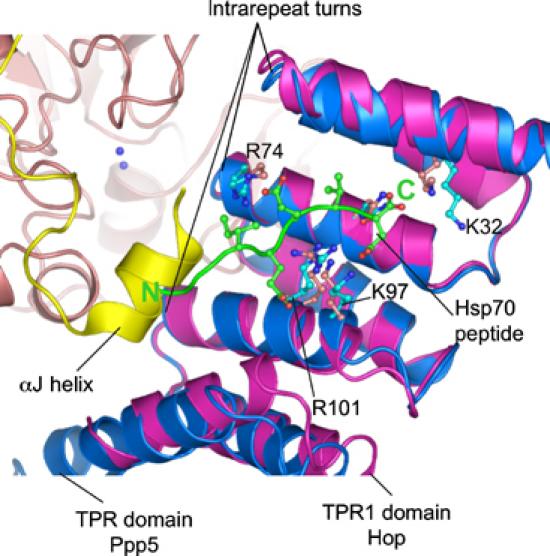
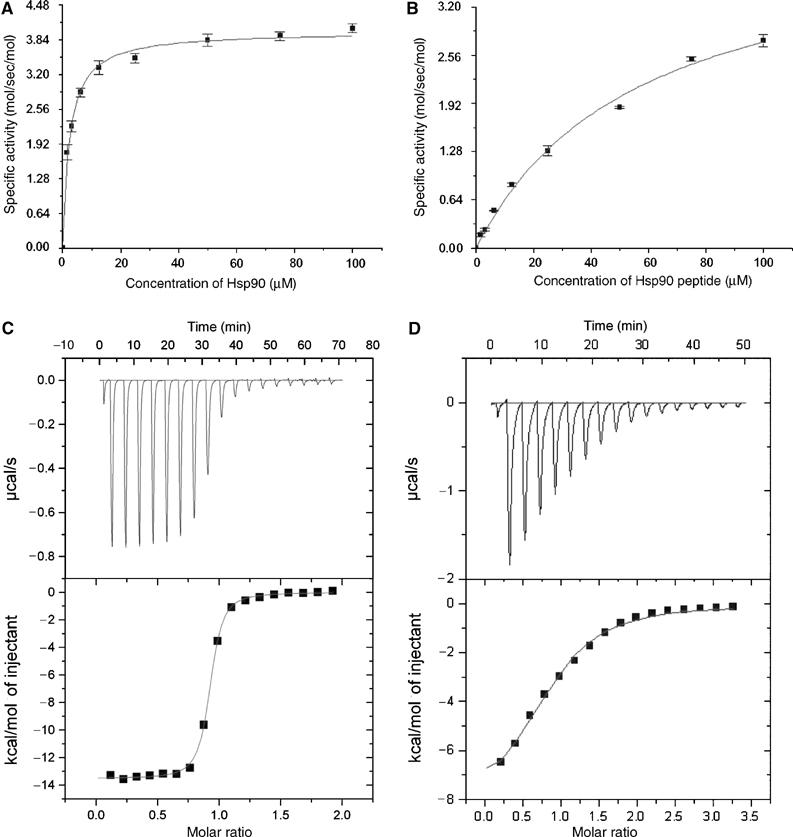
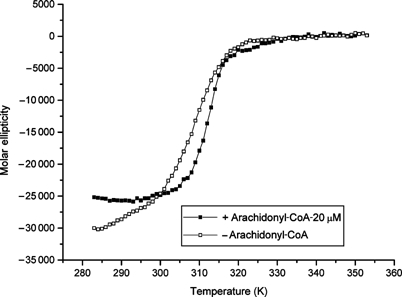
Protein phosphatase 5 (Ppp5) is a serine/threonine protein phosphatase comprising a regulatory tetratricopeptide repeat (TPR) domain N-terminal to its phosphatase domain. Ppp5 functions in signalling pathways that control cellular responses to stress, glucocorticoids and DNA damage. Its phosphatase activity is suppressed by an autoinhibited conformation maintained by the TPR domain and a C-terminal subdomain. By interacting with the TPR domain, heat shock protein 90 (Hsp90) and fatty acids including arachidonic acid stimulate phosphatase activity. Here, we describe the structure of the autoinhibited state of Ppp5, revealing mechanisms of TPR-mediated phosphatase inhibition and Hsp90- and arachidonic acid-induced stimulation of phosphatase activity. The TPR domain engages with the catalytic channel of the phosphatase domain, restricting access to the catalytic site. This autoinhibited conformation of Ppp5 is stabilised by the C-terminal alphaJ helix that contacts a region of the Hsp90-binding groove on the TPR domain. Hsp90 activates Ppp5 by disrupting TPR-phosphatase domain interactions, permitting substrate access to the constitutively active phosphatase domain, whereas arachidonic acid prompts an alternate conformation of the TPR domain, destabilising the TPR-phosphatase domain interface.
Figures
Similar articles
-
Conformational diversity in the TPR domain-mediated interaction of protein phosphatase 5 with Hsp90.Structure. 2006 Mar;14(3):415-26. doi: 10.1016/j.str.2005.12.009. Structure. 2006. PMID: 16531226
-
A novel tetratricopeptide repeat (TPR) containing PP5 serine/threonine protein phosphatase in the malaria parasite, Plasmodium falciparum.BMC Microbiol. 2001;1:31. doi: 10.1186/1471-2180-1-31. Epub 2001 Nov 28. BMC Microbiol. 2001. PMID: 11737864 Free PMC article.
-
Identification of amino acids in the tetratricopeptide repeat and C-terminal domains of protein phosphatase 5 involved in autoinhibition and lipid activation.Biochemistry. 2001 Sep 4;40(35):10485-90. doi: 10.1021/bi010999i. Biochemistry. 2001. PMID: 11523989
-
Molecular mechanisms underlying inhibition of protein phosphatases by marine toxins.Front Biosci. 1999 Oct 1;4:D646-58. doi: 10.2741/dawson. Front Biosci. 1999. PMID: 10502549 Review.
-
Structure and function of the co-chaperone protein phosphatase 5 in cancer.Cell Stress Chaperones. 2020 May;25(3):383-394. doi: 10.1007/s12192-020-01091-3. Epub 2020 Apr 2. Cell Stress Chaperones. 2020. PMID: 32239474 Free PMC article. Review.
Cited by
-
Steroid Receptor-Associated Immunophilins: A Gateway to Steroid Signalling.Clin Biochem Rev. 2015 May;36(2):31-52. Clin Biochem Rev. 2015. PMID: 26224894 Free PMC article. Review.
-
A human-based multi-gene signature enables quantitative drug repurposing for metabolic disease.Elife. 2022 Jan 17;11:e68832. doi: 10.7554/eLife.68832. Elife. 2022. PMID: 35037854 Free PMC article.
-
Characterization of protein phosphatase 5 from three lepidopteran insects: Helicoverpa armigera, Mythimna separata and Plutella xylostella.PLoS One. 2014 May 13;9(5):e97437. doi: 10.1371/journal.pone.0097437. eCollection 2014. PLoS One. 2014. PMID: 24823652 Free PMC article.
-
Advances in the structures, mechanisms and targeting of molecular chaperones.Signal Transduct Target Ther. 2025 Mar 12;10(1):84. doi: 10.1038/s41392-025-02166-2. Signal Transduct Target Ther. 2025. PMID: 40069202 Free PMC article. Review.
-
PTH stimulation of Rankl transcription is regulated by SIK2 and 3 and mediated by CRTC2 and 3 through action of protein phosphatases 1, 2, 4, and 5.J Biol Chem. 2025 Aug;301(8):110434. doi: 10.1016/j.jbc.2025.110434. Epub 2025 Jul 1. J Biol Chem. 2025. PMID: 40609791 Free PMC article.
References
-
- Andreeva AV, Kutuzov MA (1999) RdgC/PP5-related phosphatases: novel components in signal transduction. Cell Signal 11: 555–562 - PubMed
-
- Barton GJ, Cohen PTW, Barford D (1994) Conservation analysis and structure prediction of the protein serine/threonine phosphatases: sequence similarity with diadenosine tetraphosphatase from E. coli suggests homology to the protein phosphatases. Eur J Biochem 220: 225–237 - PubMed
-
- Borthwick EB, Zeke T, Prescott AR, Cohen PTW (2001) Nuclear localization of protein phosphatase 5 is dependent on the carboxy-terminal region. FEBS Lett 491: 279–284 - PubMed
-
- Brünger AT, Adams PD, Clore GM, DeLano WL, Gros P, Grosse-Kunstleve RW, Jiang JS, Kuszewski J, Nilges M, Pannu NS, Read RJ, Rice LM, Simonson T, Warren GL (1998) Crystallography and NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr D 54: 905–921 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
